 Вход
Вход Регистрация
Регистрация





Да ради бога...Пусть растут...Вместе с ВПДЕ!Если смогут...)))Так шта... смысл тренировать медленные есть.
Добро пожаловать на ЖЕЛЕЗНЫЙ ФАКТОР!
Рекомендуем Вам зарегистрироваться, чтобы получить полный доступ к форуму. После регистрации Вам будет разрешено создавать топики, писать сообщения, загружать и просматривать фотографии, оценивать посты других форумчан, управлять собственным профилем на форуме и многое другое. Личные сообщения доступны после 50 оставленных на форуме сообщений . Полный доступ к разделу "Химия" так же доступен после 50 сообщений. Если у Вас уже есть аккаунт, войдите здесь, либо зарегистрируйтесь!
Рекомендуем Вам зарегистрироваться, чтобы получить полный доступ к форуму. После регистрации Вам будет разрешено создавать топики, писать сообщения, загружать и просматривать фотографии, оценивать посты других форумчан, управлять собственным профилем на форуме и многое другое. Личные сообщения доступны после 50 оставленных на форуме сообщений . Полный доступ к разделу "Химия" так же доступен после 50 сообщений. Если у Вас уже есть аккаунт, войдите здесь, либо зарегистрируйтесь!

Думай! Научный подход в бодибилдинге

 74 голосов
74 голосов
Автор темы:
Disco
, апр 29 2009 09:38
#10741

 Отправлено 27 ноября 2012 - 11:17
Отправлено 27 ноября 2012 - 11:17
STATHAM.72
-

- Привилегированный пользователь
-

- 15 012 сообщений
Завсегдатай
-
Имя: Андрей -
Пол: Мужчина -
Город: провинциальный
"Мощно и часто,а после-декондишн"...В этом то и соль...Что разрушили сверх меры,А СИНТЕЗ после двух суток вернется к исходному!А организму это и нужно!Не нужно ничего сверх-строить!И содержать...Выстроит то же самое,а то и меньше еще...Поди куево?И содержать не надо...А будет ли новая треня-не известно!)))
 Наверх
Наверх
#10742
Отправлено 27 ноября 2012 - 11:23
Михалы4
-

- Участник форума
-
- 25 967 сообщений
body_by_mma
-
Имя: Антон -
Пол: Мужчина -
Город: Москва
-30+30)))Вот ведь мля-херь!Медленные оказывается отрубаются на 30-40 секунде и ровно за столько же восстанавливаются)))
Не пойму, с чем ты не согласен-то.
Селуянов пишет, что отрезок в 30 сек. позволяет ММВ накопить оптимус Н+ и Кр. (В условиях рабочей ишемии так называемой "стато-динамики").
Опять же, последующий за ним 30 сек. период отдыха позволяет удержать оптимус Н+ и Кр в клетке. Что, опять же, вроде бы выглядит логично.
Получаем метаболический интервальный тренинг.
Вполне себе здравая идея.
Селуянов – Гиперплазия миофибрилл в медленных мышечных волокнах
С цифрами можно, конечно, спорить, но принцип всё равно останется тем же.
Тем более что данный протокол нагрузки ничем не лучше и не хуже других, – как и все имеющиеся у человечества протоколы нагрузки, он взят с того же самого потолка – "Откуда берутся книги? Из других книг."
Сообщение изменено: Михалы4 (27 ноября 2012 - 11:35)
#10743
Отправлено 27 ноября 2012 - 11:31
STATHAM.72
-
- Привилегированный пользователь
-
- 15 012 сообщений
Завсегдатай
-
Имя: Андрей -
Пол: Мужчина -
Город: провинциальный
Да просто все эти "технические характеристики"=по ВПДЕ!Как там у Б.Джонстона?В книжке...ВПДЕ-актив=до 50 секунд!Причём тут НПДЕ???Сделайте долгую разминку...После адаптационные сеты на 15 репсов...После натяги на ВПДЕ+дропы на более чем полминуты...Медленные везде получат свои пиСты!Они вообще свое получали,как только юзер гантели в руки взял...Несколько лет назад...Они всё ещё-растут?)))Не пойму, с чем ты не согласен-то. Селуянов пишет, что отрезок в 30 сек. позволяет ММВ накопить оптимус Н+ и Кр.
#10744
Отправлено 27 ноября 2012 - 11:37
STATHAM.72
-
- Привилегированный пользователь
-
- 15 012 сообщений
Завсегдатай
-
Имя: Андрей -
Пол: Мужчина -
Город: провинциальный
И последнее...Даже если Селуянов ПРАФФ по креатину,ионам и накоплению или натягам...То вовсе нету никакой разницы по стимуляции всех ДЕ!Все едино для ВСЕХ!Ибо гипоксия!
И уж если окончательно окуеть по медленным,то сеты должны быть длинее 1 минуты!А не 30 секунд)))Именно по этому Пол Борессен и хуячил 50 репсов разом,а после натяги+дропы...
И уж если окончательно окуеть по медленным,то сеты должны быть длинее 1 минуты!А не 30 секунд)))Именно по этому Пол Борессен и хуячил 50 репсов разом,а после натяги+дропы...
#10745
Отправлено 27 ноября 2012 - 11:41
Михалы4
-
- Участник форума
-
- 25 967 сообщений
body_by_mma
-
Имя: Антон -
Пол: Мужчина -
Город: Москва
Даже если Селуянов ПРАФФ по креатину,ионам и накоплению или натягам...То вовсе нету никакой разницы по стимуляции всех ДЕ!Все едино для ВСЕХ!Ибо гипоксия!
Ну так он об этом и пишет. Именно так всё и есть. Принципы развития всех типов абсолютно одинаковы. (В условиях гипоксии).
Интервальный метаболический тренинг – это ноу-хау, применяющееся ко всем типам ДЕ, не только к медленным.
Натяги на ВПДЕ+дропы на более чем полминуты...Медленные везде получат свои пиСты!
Ноу-хау и Нобелевская премия Селуянова – это сугубо ОТДЕЛЬНЫЙ тренинг ММВ, без затрагивания быстрых! (По возможности).
С быстрыми медленные замесить любой дурачок сможет!
А ему надо "МЕДЛЕННЫХ" Олимпийских чемпионов готовить – сверхлюдей.
Ivan Drago Rules the World
Сообщение изменено: Михалы4 (27 ноября 2012 - 11:56)
#10746
Отправлено 28 ноября 2012 - 07:46
KomRADik
-

- Участник форума
-
- 1 053 сообщений
Новичок
а можно понять что такое "хорошо"? интересуют практические результаты к примеру вес, рост, процент жира, бицепс, талия, рабочие веса было->стало
Да я вроде уже где говорил об этом. В среднем, это плюс 5 кг веса в 3 месяца, 2,5 - 5 кг от травмы до травмы в жиме лежа (трени раз 2-3 недели), 5 кг в приседе (при том, что до этого был вработка, то есть далеко не новичек). Для меня это был самый большой темп роста, может кто-то растет быстрее - я не знаю. Почему считаю схему хорошей, потому что рост достаточно стабилен, можно спрогнозировать, сколько надо ориентировочно, чтобы подойти к таким-то таким-то весам и т.д.
#10747
Отправлено 28 ноября 2012 - 08:13
KomRADik
-
- Участник форума
-
- 1 053 сообщений
Новичок
До тех, пор пока с саркомера не будет снята охранная опалубка из десмина, предпринимать что-либо травматическое контр-продуктивно. Сдаётся мне, что ни окаких двух неделях там не может быть и речи. При этом необходимо помнить, что сверхкомпенсация сократительных элементов (добавление дополнительных параллельных нитей в саркомер) является КОНЕЧНОЙ фазой восстановления.
На мой взгляд, можно даже чаще. Дело в том, что как бы ты не старался, травматику равномерно по волокнам ты не положешь. Одни волона сумеешь достать, другие нет (ведь в мыщце целый спектр волокон, которые отличаются по порогу иннервации, мощности фибрилл, развитию сарко и т.д.). Те, что протравмировались, затягиваются десмином и про нагрузку на них действительно можно на время забыть. Но есть волокна, которые не достал на первой трене. Можно достать их на второй и т.д. Т.е. травматику можно накапливать. Другое дело, что каждая последующая треня - будет заметно менее эффективной в этом плане (те волокна, что проще все достать - уйдут в разнос на первой трене, а дальше остаются всё более и более устойчивые, плюс в клетке происходят сдвиги, которые защищают волокна от травматики).
Т.е. в принципе можно организовать следующий цикл - 1) накопляж травматики в мышце, 2) трени, сопровождающие восстановление.
#10749
Отправлено 28 ноября 2012 - 09:46
Davos
-

- Участник форума
-
- 918 сообщений
Новичок
-
Пол: Мужчина -
Город: Поволжье
Нашел ответ на свой вопрос. На фите предлогают так чередовать тренировки на быстрые и медленные мв.
Не пойму почему нет прав редактировать сообщение в этой теме
Либо на одной, сначала быстрые, потом медленные.Пример 1 (чередование недель)
неделя БМВ (80-90%, 6-8 повторений, взрывной стиль, отказ)
неделя ММВ (30-50%, 30-50 сек. постоянное напряжение, отказ)
неделя Восстановления (50%, 8-12 повторений, без отказа)
Пример 2 (БМВ + ММВ на одной тренировке)
неделя БМВ+ММВ
неделя Восстановления
Не пойму почему нет прав редактировать сообщение в этой теме
#10750
Отправлено 28 ноября 2012 - 10:07
KomRADik
-
- Участник форума
-
- 1 053 сообщений
Новичок
Либо на одной, сначала быстрые, потом медленные.
На мой взгляд, лучше на одной. Начинать с быстрых (при тренировке быстрых медленные тоже как ни странно получают нагрузку), потом когда быстрые уйдут в аут, добирать объем для медленных. Получается что-то вида обратной пирамиды. Почему я думаю, что лучше на одной трене - так это потому, что при работе на быстрые - нагрузка попадает и на медленные и наоборот (при работе на медленные можно немного зацепить быстрые). То есть возможны незначительные перехлесты. К тому же работа на медленные после быстрых позволит - им быстрее восстановиться (опять же - из моих предположений).
#10751
Отправлено 28 ноября 2012 - 10:30
Davos
-
- Участник форума
-
- 918 сообщений
Новичок
-
Пол: Мужчина -
Город: Поволжье
На мой взгляд, лучше на одной. Начинать с быстрых (при тренировке быстрых медленные тоже как ни странно получают нагрузку), потом когда быстрые уйдут в аут, добирать объем для медленных. Получается что-то вида обратной пирамиды. Почему я думаю, что лучше на одной трене - так это потому, что при работе на быстрые - нагрузка попадает и на медленные и наоборот (при работе на медленные можно немного зацепить быстрые). То есть возможны незначительные перехлесты. К тому же работа на медленные после быстрых позволит - им быстрее восстановиться (опять же - из моих предположений).
Да, но тогда возникают сложности при тренировке по фулбоди, придется делать по два упражнения на каждую мг, при чем сначала все на бмв, затем на ммв.
Кстати, как вариант (при фулбоди), можно все на 1й неделе: пон - БМВ, вт -ММВ, ср - восстановление.
Сообщение изменено: Davos (28 ноября 2012 - 10:38)
#10752
Отправлено 28 ноября 2012 - 11:17
Михалы4
-
- Участник форума
-
- 25 967 сообщений
body_by_mma
-
Имя: Антон -
Пол: Мужчина -
Город: Москва
Имеет ли смысл выделять отдельный день для специализации на ММВ?
Если хочешь стать "МЕДЛЕННЫМ" Олимпийским чемпионом – тогда можно попробовать тренировать только ММВ. (А быстрые не тренировать вообще).
Во всех других случаях весь пул (и быстрые, и медленные) прокачивается разом.
Придется делать по два упражнения на каждую МГ, при чем сначала все на БМВ, затем на ММВ.
Никаких отдельных замесов на быстрые и медленные делать не нужно. Тройной дропсет прокачивает весь пул разом и равномерно.
пон - БМВ, вт -ММВ
Владимир Кравцов так делает. Разбивает тренировку всего пула на две части – БМВ и ММВ. Микроцикл у него 12 дней. (По ссылке, правда, несколько другой вариант. Про тренинг конкретно жимовых в режиме ММВ он говорил в одном из интервью).
Сообщение изменено: Михалы4 (28 ноября 2012 - 11:50)
#10753
Отправлено 28 ноября 2012 - 12:17
Davos
-
- Участник форума
-
- 918 сообщений
Новичок
-
Пол: Мужчина -
Город: Поволжье
Владимир Кравцов так делает.
В этой программе тренировка ММВ у него почему-то только на ноги и спину.
#10755
Отправлено 28 ноября 2012 - 03:28
KomRADik
-
- Участник форума
-
- 1 053 сообщений
Новичок
Вопроса не понял. Адаптация к травматике в любом случае будет и будет довольно быстрой. Для этого и нужна "нетравматика". Мы с одной стороны - поддерживаем ЭОС, даем натяг (90% ПМ, как я делал - это всё же натяг), даём постоянную иннервацию на волокна (а я где-то у кого-то в работах видел мысли о том, что новые ядра без иннервации могут не присоединяться или не удерживаться, там какая-то схема регуляции через Кальциневрин) и заодно отрабатываем нервно-мышечные связи (да и рецепторы гольджи блокируем), ну и ждем пока "опалубка" из десмина отойдет.
#10756
Отправлено 29 ноября 2012 - 02:30
ctjan
-

- Участник форума
-
- 6 453 сообщений
Бывалый
-
Пол: Не определился -
Город: где то рядом
Не нужно никакого немедленного поддержания травматических тренировок, тренировок сопровождающих восстановление. Т.к. если стресс стимул был оптимальный то, синтез после сделает свое дело с свехкомпенсацией, поэтому боятся, что восстановятся белковые структуры только до исходного уровня нечего. Т.к. работает адаптационный механизм. И лезть в этот процесс не стоит, а наоборот помочь организму выполнить свою адаптационную программу правильным питанием и отдыхом. Вопрос в том, чтобы подобрать оптимальный стресс стимул.
#10757
Отправлено 29 ноября 2012 - 01:35
sandwolf77
-

- Модератор
-
- 21 009 сообщений
crusader of philosophy
-
Имя: Алексей -
Пол: Мужчина -
Город: Тюмень
меня очень заинтересовала!но какой интервал должен бы разделять травматику многоповторную и травматику синглов?и какая поддержка была бы оптимальна на стадии сверхкомпенсации - отдых, умеренно-тяжелая малоповторка, памп? в рамках данной альтернативы,разумеется.До тех, пор пока с саркомера не будет снята охранная опалубка из десмина, предпринимать что-либо травматическое контр-продуктивно.
Сдаётся мне, что ни окаких двух неделях там не может быть и речи.
При этом необходимо помнить, что сверхкомпенсация сократительных элементов (добавление дополнительных параллельных нитей в саркомер) является КОНЕЧНОЙ фазой восстановления.
Гликолиз, репсы...
А мне вот почему-то кажется, что для наращивания дополнительных миофиламентов первостепенен механостимул. Типа вот этого:
Говорят, у Кубика на этом система построена. "Смертельные сеты" (травматическая высокоповторка) + синглы.
Ну, я как бы не настаиваю. Просто напомнил альтернативу. Может кого заинтересует.

#10758
Отправлено 29 ноября 2012 - 09:00
roma2
-

- Привилегированный пользователь
-
- 9 032 сообщений
Чатланин Уэф
-
Имя: Уэф -
Пол: Мужчина -
Город: г. "Трех шурупов", Планета Плюк, 215 в Тентуре, галактика Кин-дза-дза в Спирали
Глянь на табличку YUNa! Там идет период травматических трений с последующим длительным отдыхом.До тех, пор пока с саркомера не будет снята охранная опалубка из десмина, предпринимать что-либо травматическое контр-продуктивно.
#10759
Отправлено 29 ноября 2012 - 09:21
Goldberg
-

- Участник форума
-
- 1 908 сообщений
Fear of the dark
-
Имя: Алексей -
Пол: Мужчина -
Город: Донецк
3-4 недели Вишняк советует делать базовые упры с субмаксимальными весами.Пример 1 (чередование недель) неделя БМВ (80-90%, 6-8 повторений, взрывной стиль, отказ) неделя ММВ (30-50%, 30-50 сек. постоянное напряжение, отказ) неделя Восстановления (50%, 8-12 повторений, без отказа)
#10760
Отправлено 30 ноября 2012 - 12:24
Deejgrey
-

- Участник форума
-
- 182 сообщений
Новичок
-
Имя: Сергей -
Пол: Мужчина -
Город: Евпатория
К вопросу о механическом повреждении мышц.
Фрайден Ж., Лиебер Р. Структурное и механическое основание повреждения мышцы посредством упражнения
Friden, J. Structural and mechanical basis of exercise-induced muscle injury / J. Frieden, R.L. Lieber //Med. Sci. Sports Exerc., 1992.– Vol. 24.– No. 5.– P. 521-530.
ABSTRACT
В опытах на животных и человеке доказано, что непривычные или эксцентрические упражнения приводят к повреждению сократительной части мышечного волокна или его цитоскелета. Чаще всего эти повреждения включают: «вытекание» Z-полосы и ее «растворение», разрушение A-полосы, распад промежуточной системы филаментов, и некоаксиальность миофибрилл. Механический характер этих повреждений свидетельствует о том, что мышечные волокна повреждаются скорее из-за уровня натяжения волокна, а не абсолютного напряжения, приложенного к волокну. Мы предполагаем, что повреждение, вызванное эксцентрическими сокращениями, происходит во время выполнения упражнений, то есть в течение первых нескольких минут. Структурные повреждения больше в быстрых мышечных волокнах. В заключительной части этой статьи, мы выдвигаем гипотезу о схеме повреждения, основанной на окислительной способности мышцы, как детерминирующем факторе.
Ключевые слова: мышечное волокно, цитоскелет, повреждение миофибрилл, быстрые мышечные волокна
Напряженное упражнение вызывает изменения в гомеостазе мышцы. Хотя ткань мышцы чрезвычайно пластичная (43), разрушительные изменения в тонкой структуре мышцы могут произойти в ответ на необычные требования (17). В опытах на животных (3,28,30,31,37) и человеке (9-13,40) получены структурные повреждения после различных типов упражнения. Описанные повреждения вовлекают: 1) первичное или вторичное разрушение сарколеммы (2,6,21), раздутие или разрушение саркотубулярной системы (2), искажение сократительных компонентов миофибрилл (3,9,11-13,22,31,40), 4) повреждение цитоскелета (10, 5) повреждения внеклеточной матрицы (50).
Такие патологические изменения могут также быть замечены у людей после спринтерского бега или после бега на длинные дистанции (28,41,56), а также после силовой тренировки (14). Так как многие из этих действий содержат большую эксцентрическую составляющую сокращения и потому, что эксцентрическое упражнение, как было установлено – самая разрушительная форма упражнения, много исследователей осуществляли эксцентрические модели сокращения мышцы, чтобы изучить адаптацию к интенсивной физической тренировке (3,12,31,40).
Цель этого обзора состоит в том, чтобы описать реакцию миофибрилл и цитоскелета на высокоинтенсивное упражнение, которое производит повреждение в архитектуре мышцы. Кроме того, мы рассматриваем механику эксцентричных сокращений и выдвигаем гипотезу о механизме, посредством которого вызывается морфологическое повреждение мышечного волокна.
Многие из структурных отклонений после эксцентрического сокращения затрагивают Z-диск. Точная тонкая структура Z-диска полностью не изучена. Однако, так как мы рассматриваем структурные свойства Z-диска в интерпретации миофибриллярных повреждений посредством упражнения, мы чувствуем необходимость дать краткий обзор исследований, посвященных ультраструктуре Z-диска.
Ультраструктура Z-диска. В поперечном сечении, Z-диск появляется как тканная корзина или квадратная решетка. Сообщается, что образцы обмениваются в течение сокращения (29). Чтобы объяснить наблюдаемую ультраструктуру были представлены различные модели (8,15,23,25-27,29,35,47-49,54). Первый структурный анализ Z-диска позвоночных (лягушка m.semitendinosus) был представлен Knappeis и Carlsen (27). Они предположили, что каждая тонкая нить I-полосы с одной стороны Z-диска помещена в центре и равноудалена от четырех тонких филаментов с противоположной стороны диска, и также связана с четырьмя филаментами Z-диска. Franzini-Armstrong и Porter (
 предположили, что Z-диск – это мембрана, к которой прикрепляются концы филаментов I-диска. Reedy (47) описал решетку, состоящую из четырех раскручивающихся жил, подходящих к Z-диску от каждого I-филамента. Kelly (25) предположил, что Z-диск сформирован «жилами», происходящими от одного саркомера, и перекрученными с «жилами» с противоположного саркомера, и возвращающимися, чтобы присоединиться к I-нитям на своей стороне. Каждая I-нить раскололась бы на две «жилы». Rowe (49) также предложил модель, основанную на перекрученных «жилах», хотя он предполагал, что перекрученные «жилы» за счет шпильки присоединились к смежной I-филаменте на своей стороне, но без соединения с «жилами» с противоположной стороны. Kelly и Cahill (26) предложили модель в которой каждая I-нить, дает начало четырем изгибающимся нитям Z-диска, которые простираются на I-филаменты противоположного саркомера. Ullrick с соавт. (54) предположил, что один тонкий филамент, входящий в Z-диск непрерывно связан с тремя изогнутыми Z-филаментами, которые объединяют это с тремя другими тонкими филаментами того же самого саркомера. В более позднем исследовании, Goldstein с соавт. (15) показал, что Z-диск мог бы состоять из концов актиновых филаментов, организованных в комплекс гребенчатой формы, связанных косыми перемычками филаментов. В дополнение ко всем этим разнообразным теориям, нужно также учитывать, что различия в описаниях могли произойти из-за разновидностей и/или различий типов волокон, так же как и технических изменений, типа различных процедур фиксации, толщины сечения и косого направления сечения.
предположили, что Z-диск – это мембрана, к которой прикрепляются концы филаментов I-диска. Reedy (47) описал решетку, состоящую из четырех раскручивающихся жил, подходящих к Z-диску от каждого I-филамента. Kelly (25) предположил, что Z-диск сформирован «жилами», происходящими от одного саркомера, и перекрученными с «жилами» с противоположного саркомера, и возвращающимися, чтобы присоединиться к I-нитям на своей стороне. Каждая I-нить раскололась бы на две «жилы». Rowe (49) также предложил модель, основанную на перекрученных «жилах», хотя он предполагал, что перекрученные «жилы» за счет шпильки присоединились к смежной I-филаменте на своей стороне, но без соединения с «жилами» с противоположной стороны. Kelly и Cahill (26) предложили модель в которой каждая I-нить, дает начало четырем изгибающимся нитям Z-диска, которые простираются на I-филаменты противоположного саркомера. Ullrick с соавт. (54) предположил, что один тонкий филамент, входящий в Z-диск непрерывно связан с тремя изогнутыми Z-филаментами, которые объединяют это с тремя другими тонкими филаментами того же самого саркомера. В более позднем исследовании, Goldstein с соавт. (15) показал, что Z-диск мог бы состоять из концов актиновых филаментов, организованных в комплекс гребенчатой формы, связанных косыми перемычками филаментов. В дополнение ко всем этим разнообразным теориям, нужно также учитывать, что различия в описаниях могли произойти из-за разновидностей и/или различий типов волокон, так же как и технических изменений, типа различных процедур фиксации, толщины сечения и косого направления сечения.Maruyama и др. (36) считает невозможным представить, как толстые филаменты могли сохранить присутствие в саркомере, не предполагая, что их свободные концы были закреплены к Z-диску некоторой непрерывной упругой структурой. Это также гарантировало бы, что, после сверхрастяжения (то есть, когда наложение актина и миозина исчезает), восстановление мышечными волокнами своей начальной длины и сократительных свойств. Эти функции могли быть приписаны существованию титина, соединенного с толстыми филаментами посредством актиновых филаментов и/или Z-диска (19,36, рис. 1). Согласно этим авторам, возможно, что титиновые филаменты, происходящие из филаментов миозина также могут связаться с другим упругим компонентом, типа линии N2. находящейся близко к Z-диску, где расположен небулин. Другими словами, эти два белка с очень высокой молекулярной массой, очевидно, играют существенную роль в обеспечении осевой непрерывности для производства напряжения покоя, сопротивления растяжению, и обеспечения целостности Z-диска (18,36,44). Кроме того, белки цитоскелета: десмин, виментин, и синемин найдены на периферических Z-дисках миофибрилл, указывая, что эти белки нужно считать интегральными саркомерными компонентами, важными в структуре мышцы и ее механике (45).
Разрушение миофибрилл после эксцентрического сокращения. Несколько авторов наблюдали изменения в миофибриллах после повторных эксцентричных сокращений (3,11-13,28,40). Friden с соавт. (12) идентифицировали дезорганизацию Z-диска миофибрилл в m. soleus человека через 3 дня после бега по наклонной плоскости. Эти авторы установили характерный вид изменений в сократительном аппарате, включая: расширение, смазывание, или даже общее миофибриллярное разрушение на уровне Z-дисков. Z-полосы смежных миофибрилл не соответствовали друг другу и были загзагообразными (рис. 2A). Дополнительно, некоторые A-полосы в пределах поврежденного саркомера не соответствовали друг другу (рис. 2B) и в некоторых случаях полностью отсутствовали толстые филаменты (9,13). Armstrong с соавт. (3) нашел, что у крыс сразу же после бега по наклонной плоскости, в волокнах медленно разгибающихся мышц были обнаружены разрушения, которые были видны и оптический и электронный микроскоп. Ogilvie с соавт. (42) сообщил о трех типах повреждений: 1) центральные разрушения A-полосы, 2) роспуск Z-линии, и 3) сгущенные волокна. Ogilvie и сотрудники сообщили, что 90 % изменений были связаны с разрушениями A-полосы. Newham с соавт. (40) показал, что, в биопсиях, полученных из m. quadriceps femoris человека, выполняющего эксцентрические сокращения, происходят обширные разрушения саркомеров. Собранные все вместе, эти авторы представили ясное свидетельство драматических структурных отклонений после эксцентрического упражнения.
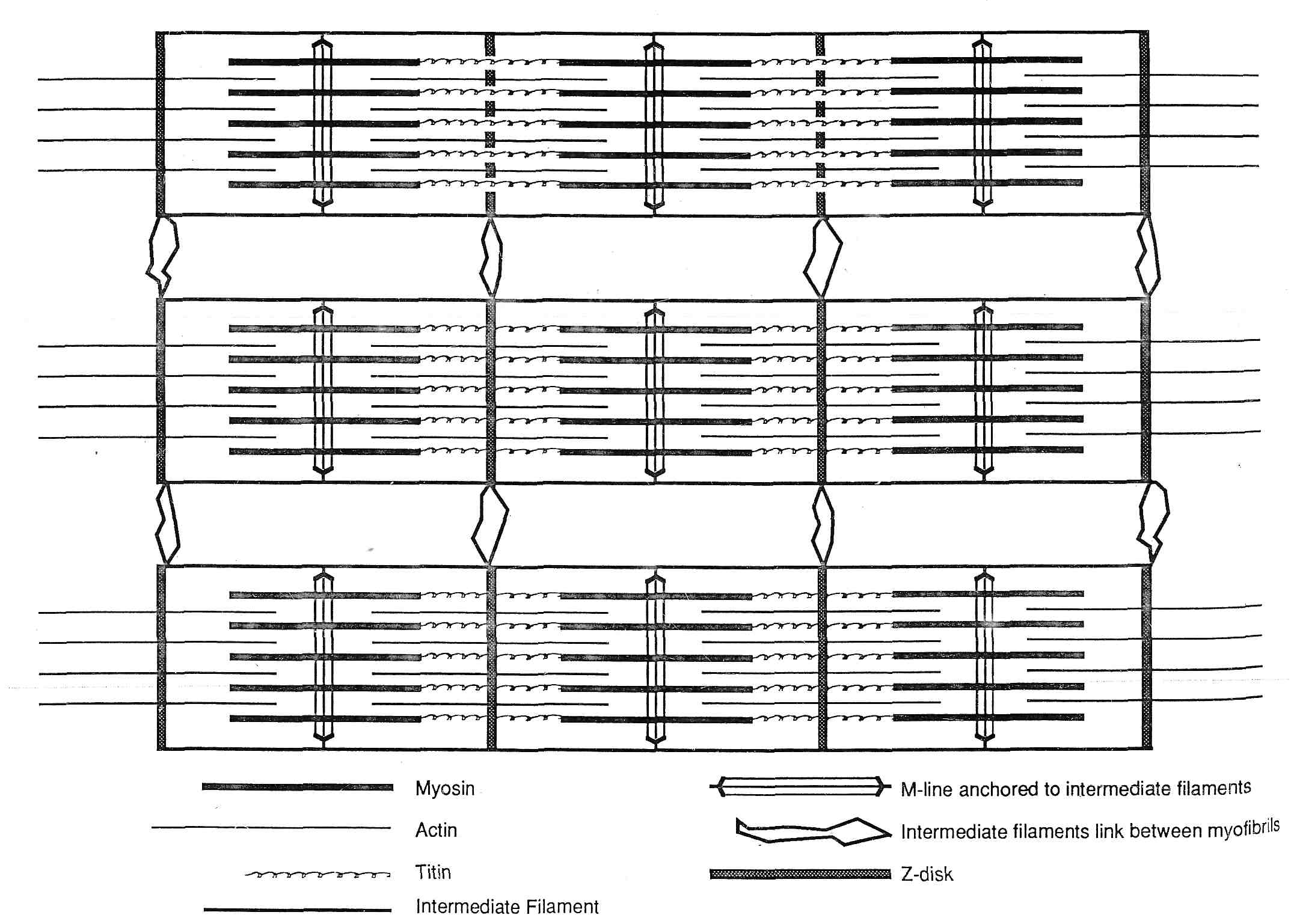
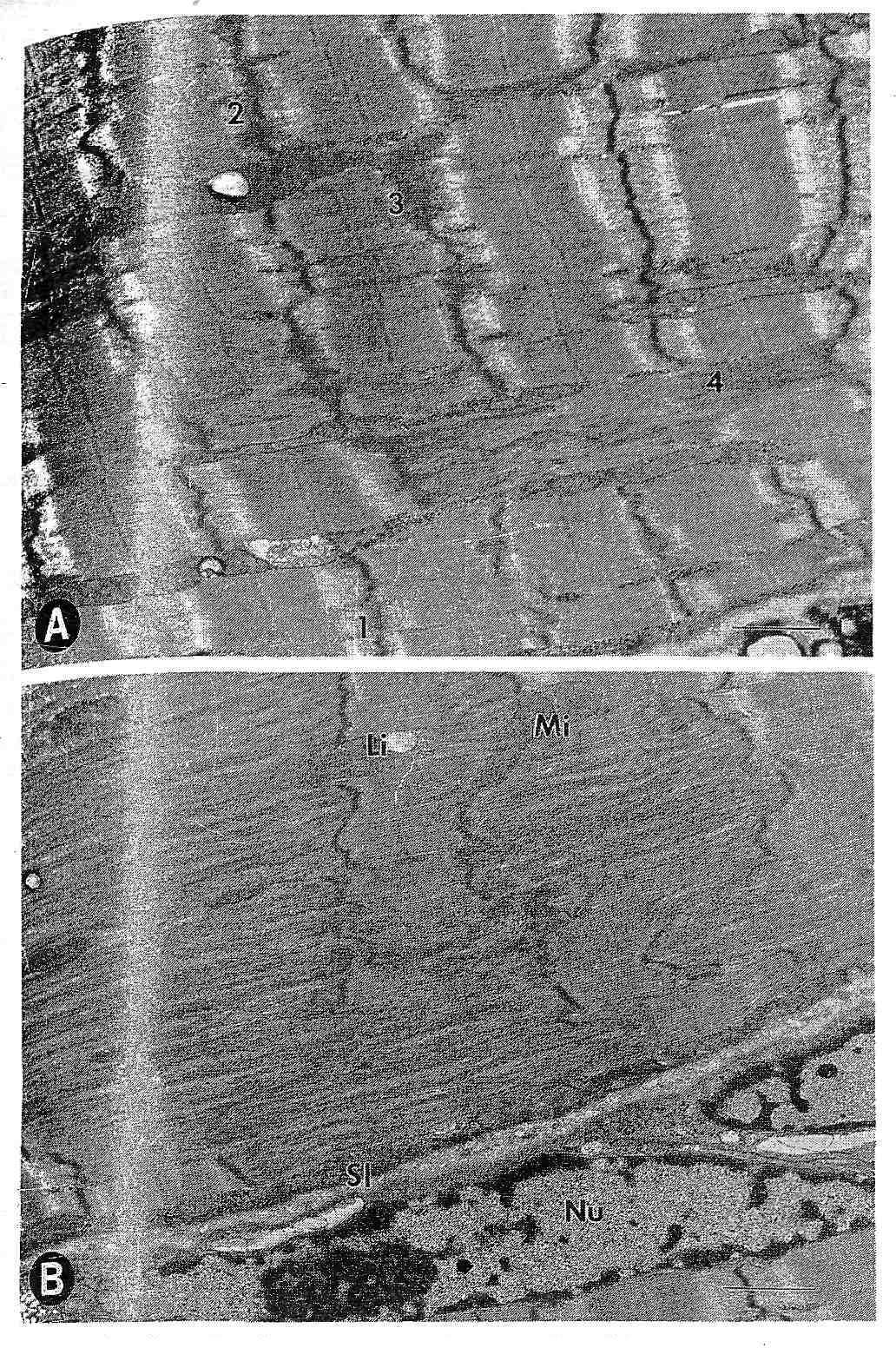
Рис.1. Схематическая диаграмма расположения саркомерных сократительных белков и белков цитоскелета.
Биологическое значение разрушения Z-диска, возникающего во время эксцентрических сокращений до настоящего времени неизвестно. Это, однако, является общим нарушением в мышце (5). Fischman с соавт. (7) и Meltzer с соавт. (38) сообщили о «размытом» Z-диске у контрольной группы. Z-дисковое вытекание, кажется, является первичным миофибриллярный ответом на измененные физические и метаболические ситуации. Ясная связь между Z-дисковым вытеканием и высокоинтенсивным физическим упражнением демонстрировалась множеством исследователей (3,12,40). Состав неорганизованного Z-дискового материала полностью не известен, но связывающие белки Z-диска десмин, и спектрин, как было показано посредством электронного микроскопа, могут быть элементами плотного волокнистого материала в областях «размытого» Z-диска (10,52). Являются ли эти полученные данные проявлением саркомерогенезиса, дегенеративных событий, или неустойчивости в обмене миофибриллярных белков не известно.
Рис.2. Продольная электронная микрофотография мышцы vastus lateralis человека после интенсивного эксцентрического упражнения. (A) Центральное разрушение Z-полосы, демонстрирующей различные степени вытекания. 1 = появление волнистой Z-полосы, в виде зигзага, 2 = умеренное вытекание Z-полосы, 3 = более серьезное повреждение Z-полосы: вытекание затрагивает часть I-диска, 4 = разрушение Z-полосы и распад миофибриллярных компонентов во всем саркомере. ( более серьезное разрушение Z-полосы (смазывание) и центральное разрушение области A-полосы сократительного аппарата. Li = капелька липида, Mi = митохондрии, Sl = sarcolemma, и NU = ядро, масштаб = 1 мкм.
более серьезное разрушение Z-полосы (смазывание) и центральное разрушение области A-полосы сократительного аппарата. Li = капелька липида, Mi = митохондрии, Sl = sarcolemma, и NU = ядро, масштаб = 1 мкм.
При использовании иммунофлюоресценции для локализации десмина в мышце vastus lateralis у человека, выполняющего эксцентрические упражнения, мы наблюдали продольные расширения флуоресцентного материала между последовательными Z-дисками в биопсиях через три дня после упражнения (10). Кроме того, эти продольные расширения были беспорядочно распределены всюду по глубине рассечения. Поперечные рассечения укрепили полученные данные, показывая частые сильно флуоресцентные пятна, вкрапленные в регулярные межфибриллярные сети. Эти изменения цитоскелета могут быть вызваны непосредственно высокой напряженностью упражнения, таким образом, представляя повреждение; или они могут представлять вторичный ответ на обширные миофибриллярные повреждения и поэтому представить саркомерогенезис. Последняя интерпретация, таким образом, указывала бы, что увеличенный синтез десмина и перестройка системы цитоскелета, является важной частью в перестройке измененного миофибриллярного материала. Десминовые филаменты, таким образом, действовали бы как механические интеграторы для ремонта саркомеров. Нам нравится эта последняя гипотеза расширенного обмена миофибриллярных и цитоскелетных белков после их деградации. Это – разумное рассмотрение факта, что копии промежуточных филаментов, окружающих Z-диск могут внести свой вклад в дополнение новых саркомеров в пределах поврежденных миофибрилл (55). Увеличение интенсивности окрашивания десмина через три дня после упражнения по сравнению с тем, что было сразу после упражнения, поддерживает это утверждение.
В дополнение к миофибриллярным и цитоскелетным повреждениям, через три дня после упражнения под ультрафиолетом было найдено увеличенное количество липофуксина. Липофуксин вообще расценивается как трудно перевариваемый остаток лизосомальной деградации (51). Факт, что ультраструктурное разрушение достигает пиковых значений через 2-3 дня после упражнения, может указывать на то, что начальное повреждение ухудшается выходом и деятельностью гидролитических ферментов в пределах волокна, далее ухудшая сократительные и цитоскелетные компоненты.
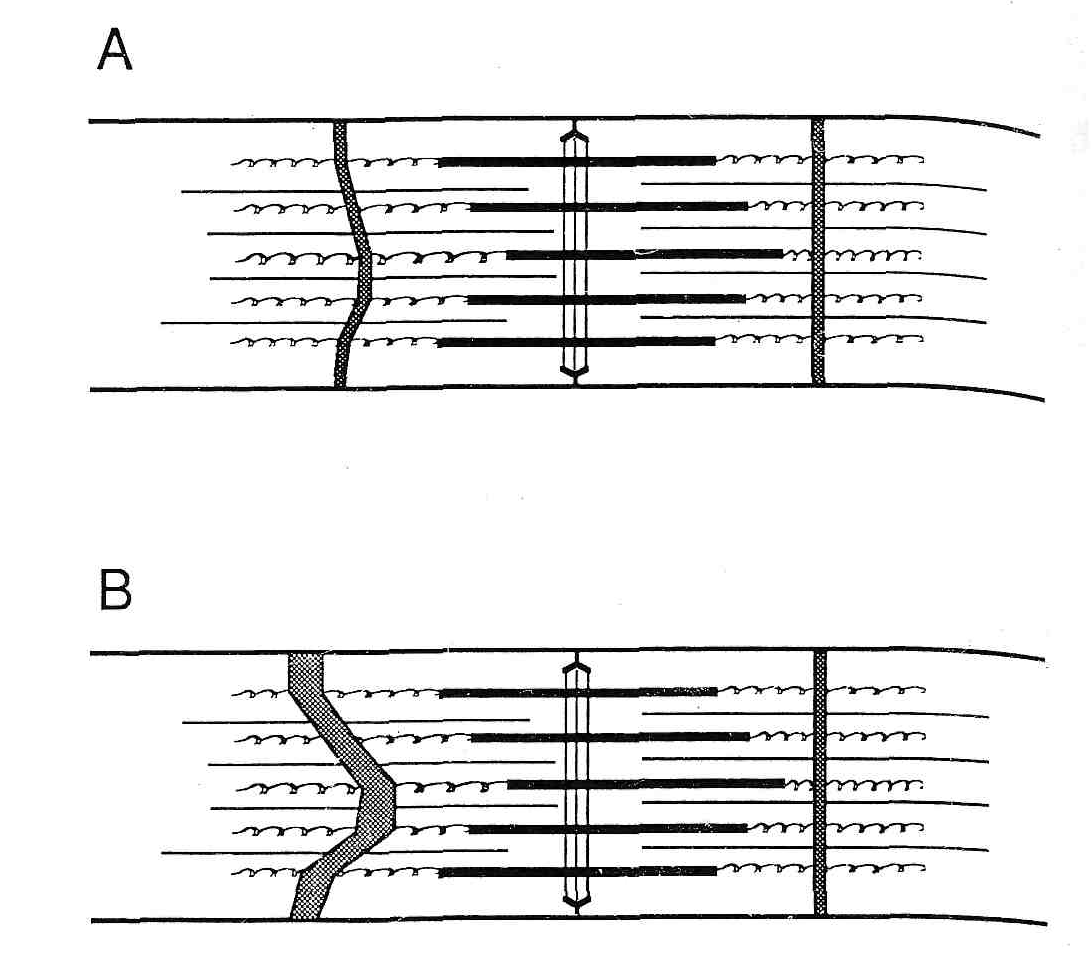
Основываясь на этих наблюдениях, можно предположить, что высокоинтенсивное упражнение может растянуть или разорвать промежуточные филаменты, связывающие Z-диск с Z-диском, а также разрушает дублирующие промежуточные филаменты Z-дискового кольца. Это разрушение молекул белка вызывает разрушение белков Z-дискового кольца, также глобулярных белков и активизирует активность лизосомальных ферментов, вызывая дальнейшую деградацию и в конечном счете разрушение Z-диска. Однако какие обстоятельства приводят к центральному или общему миофибриллярному распаду объяснить невозможно. Чтобы объяснять адаптацию цитоскелета, должно быть предпринято более полное исследование реакции внутрисаркомерного цитоскелета, что позволило бы включить в анализ оценку важности филаментов титина и небулина для того, чтобы предотвратить сверхрастяжение миофибриллы. Если бы, действительно, эти филаменты были бы физически повреждены в результате высокоинтенсивного эксцентрического упражнения, конечно, появилась бы осевая некоаксиальность толстых филаментов (18, рис. 3).
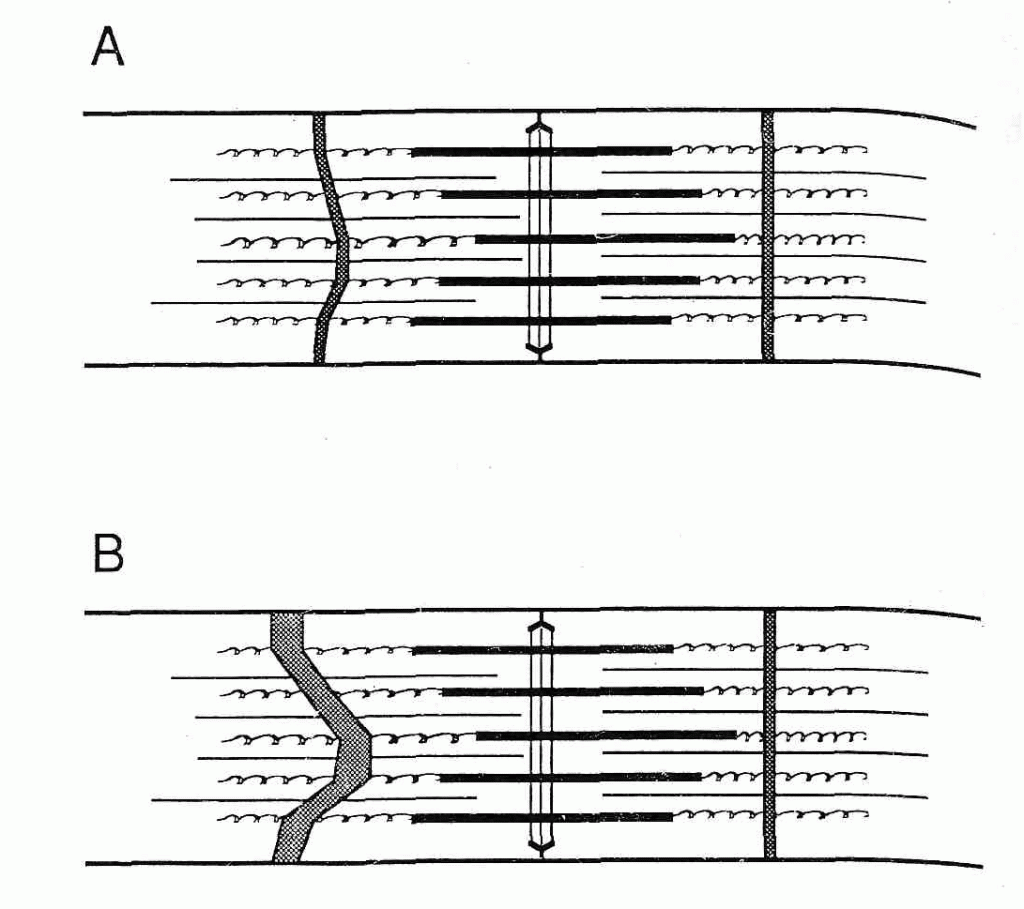
Рис.3 Схематическая диаграмма гипотетического механизма, посредством которого разрушается Z-полоса. Филаменты миозина, которые смещены из-за неравенства сил поперек саркомера, могут тянуть саркомер в одну сторону. В течение сокращения, активное смещение миозина может также сместить актин, который закреплен к Z-линии. (A) Минимальное «вытекание». ( более серьезное «размытие» Z-полосы.
Следующее начальное повреждение, связанное с дальнейшей деградацией имеет место в течение следующих часов. Этот процесс, вероятно, инициируется увеличенным количеством внутриклеточного кальция (из-за вызванной утомлением неспособности митохондрий противостоять увеличению кальция), что приводит к активации активизированных кальцием нейтральных протеаз, лизосомных протеаз, и других клеточных процессов, которые связаны с кальцием (1). Интересно, что лаборатория Армстронга недавно предоставила прямое доказательство увеличенной митохондриальной концентрации кальция в крысах, выполняющих длительный бег по наклонной плоскости. Они также показали, что агенты, которые связывали кальций (например, chelating агент, EDTA) уменьшали величину повреждения (6). Armstrong (2) также представил обзор, который подчеркивает критическую роль внутриклеточного регулирования кальция в повреждении мышцы. Мы полагаем, что более серьезное миофибриллярное повреждение, отмечаемое через два и три дня после эксцентрического упражнения, может быть результатом активизированной кальцием, увеличенной протеолитической деятельности ферментов.
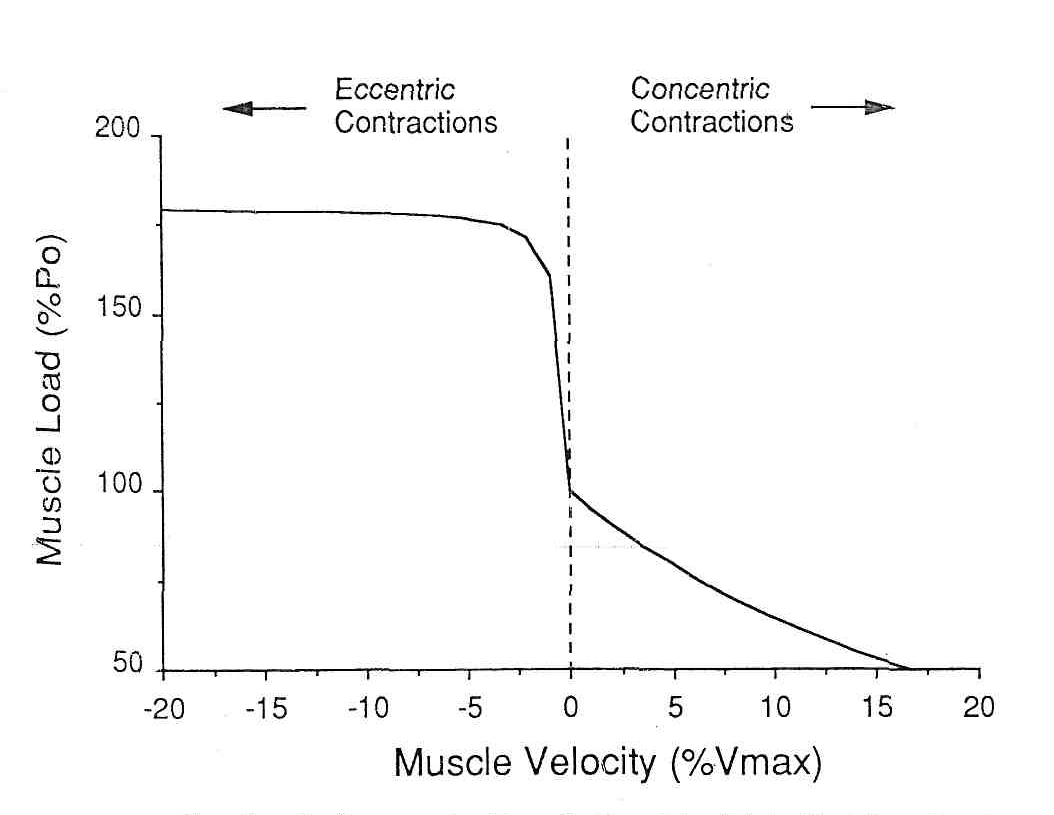
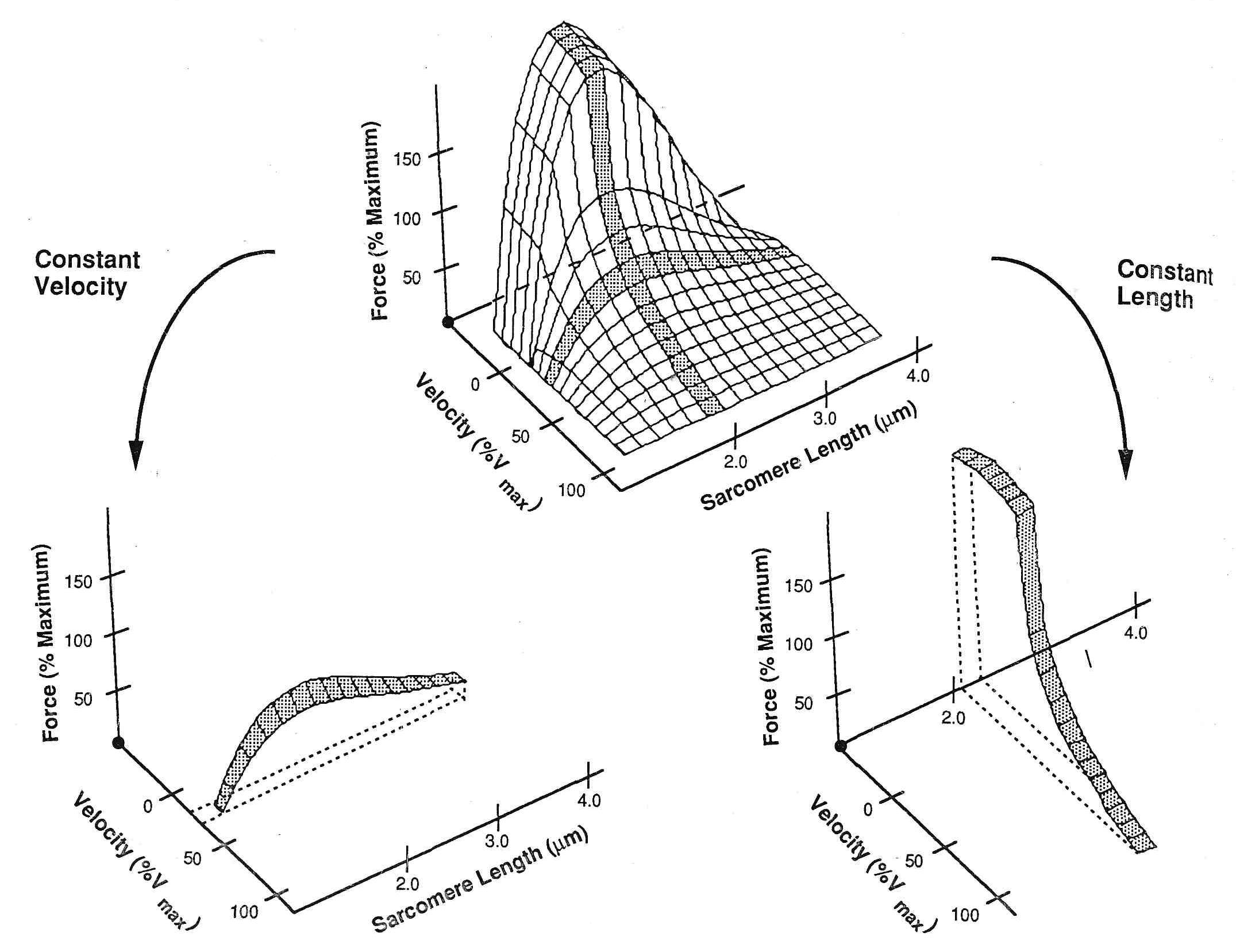
Механическое рассмотрение эксцентрических сокращений. Давно известно, что мышцы, которые удлиняются, могут противодействовать очень большим нагрузкам, и поэтому такие удлиняющие сокращения вызывают повреждение мышцы. Katz (24) объяснил, что механическим основанием этого наблюдения является зависимость «сила-скорость» Он показал, что когда мышце позволяли сокращаться, ее сила уменьшалась почти линейно (по крайней мере в областях низких скоростей) в пределах приблизительно (4%Pо х %Vmax), рис. 4. Таким образом, мышца при сокращении на 5%-ов от Vmax, уменьшает свою силу примерно на 20% от Ро. Однако, для удлинения мышцы, отношения были весьма более сложными. Удлинение мышцы только на 1% от Vmax может увеличить силу на 50% от Ро.

Рис.4 – Классическая зависимость «скорость–сила». Отметьте, что при сокращении (скорость, больше чем ноль) падение силы относительно медленное с возрастанием скорости. Однако, при удлинении мышцы (скорость сокращения меньше нуля) сила возрастает очень резко. Это может вызывать нестабильность между соседними саркомерами (c.f., рис. 3).
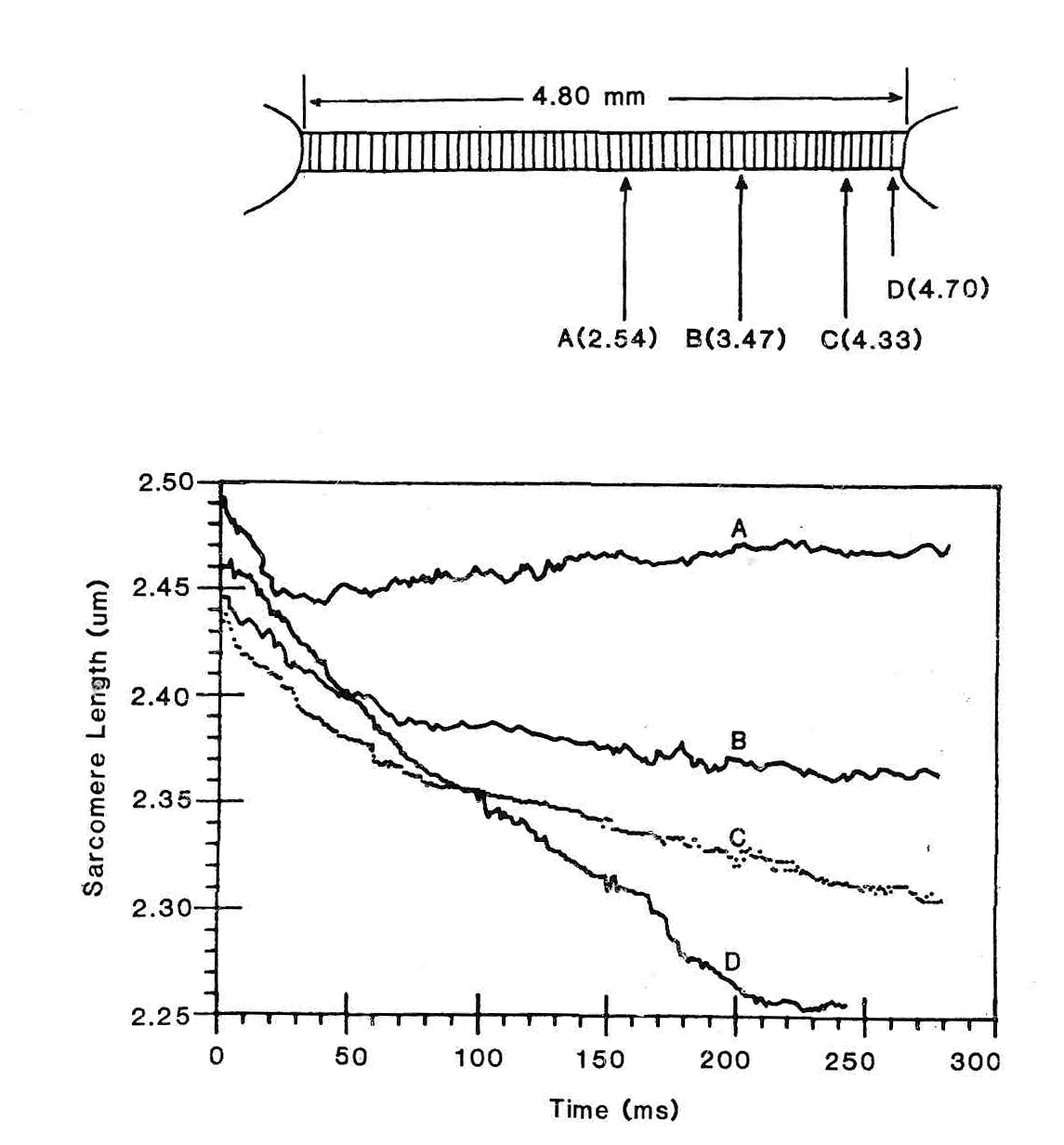
Фактически, неоднородность зависимости «сила-скорость» около нуля приводит к тому, что сила увеличивается приблизительно в 10 раз быстрее при удлинении, чем при такой же скорости сокращения. Эта неоднородность является основанием различий между соседними саркомерами с точки зрения сил, с которыми они взаимодействуют. Вспомните, что длина саркомера относительно длины мышечного волокна может измениться примерно до 1 % (20). В течение сокращений саркомера с одним закрепленным концом установлено, что его скорость для большинства саркомеров, находящихся вдоль длины мышечного волокна варьирует от 1 до 5 % от Vmax (16). Это неравенство длины и скорости может быть ясно замечено на рис.5, который представляет длину саркомера как функцию времени в различных областях изолированного волокна мышцы лягушки (30). Отметьте, что на концах мышечного волокна, саркомеры более короткие и сокращаются с высокой скоростью. К центру мышечного волокна длина саркомеров увеличивается, и скорость сокращения увеличивается. Наконец, в центре волокна, саркомеры относительно длинны и фактически медленно сокращаются в течение изометрического сокращения. Таким образом, каждая смежная популяция саркомеров вдоль волокна имеет немного различную длину и скорость, но производит одно и то же напряжение. Они производят одно и то же напряжение в силу зависимости «сила-длина» мышцы.
Рис.5. Сверху. Области вдоль мышечного волокна, где измерялась длина саркомеров. Внизу длина саркомеров вдоль изолированного волокна мышцы в течение изометрического сокращения. В этом эксперименте, концы изолированной semitendinosus лягушки были фиксированы. Мышца была активизирована, после чего измерялась длина саркомеров, расположенных вдоль длины волокна на основе лазерной дифракции. Отметьте, что саркомеры, находящиеся около конца волокна (С и D) имеют более короткую длину и более высокую скорость сокращения, тогда как саркомеры находящиеся в середине волокна (A) фактически удлиняются на медленной скорости. Поскольку эти саркомеры более длины и таким образом имеют более низкий изометрический потенциал силы, сила по длине волокна постоянна (30).
Общую зависимость «длина-сила-скорость» иллюстрирует поверхность (рис.6), в которой длина и/или скорость могут быть различны по почти бесконечному числу комбинаций, чтобы достигнуть специфической силы. Традиционные зависимости «длина-сила» и «сила-скорость» – просто частные случаи этой поверхности, когда любая длина (для зависимости «сила-скорость») или скорость (для зависимости «длина-сила») считается постоянной.
Рис.6. Общая зависимость между длиной мышцы, силой и скоростью Заметьте, что достижение определенной силы возможно при различной длине саркомера и скорости Если эту общую зависимость рассматривать при постоянной длине, получается зависимость сила-скорость (справа), если рассматривать при постоянной скорости – то получается зависимость длина –сила (справа).
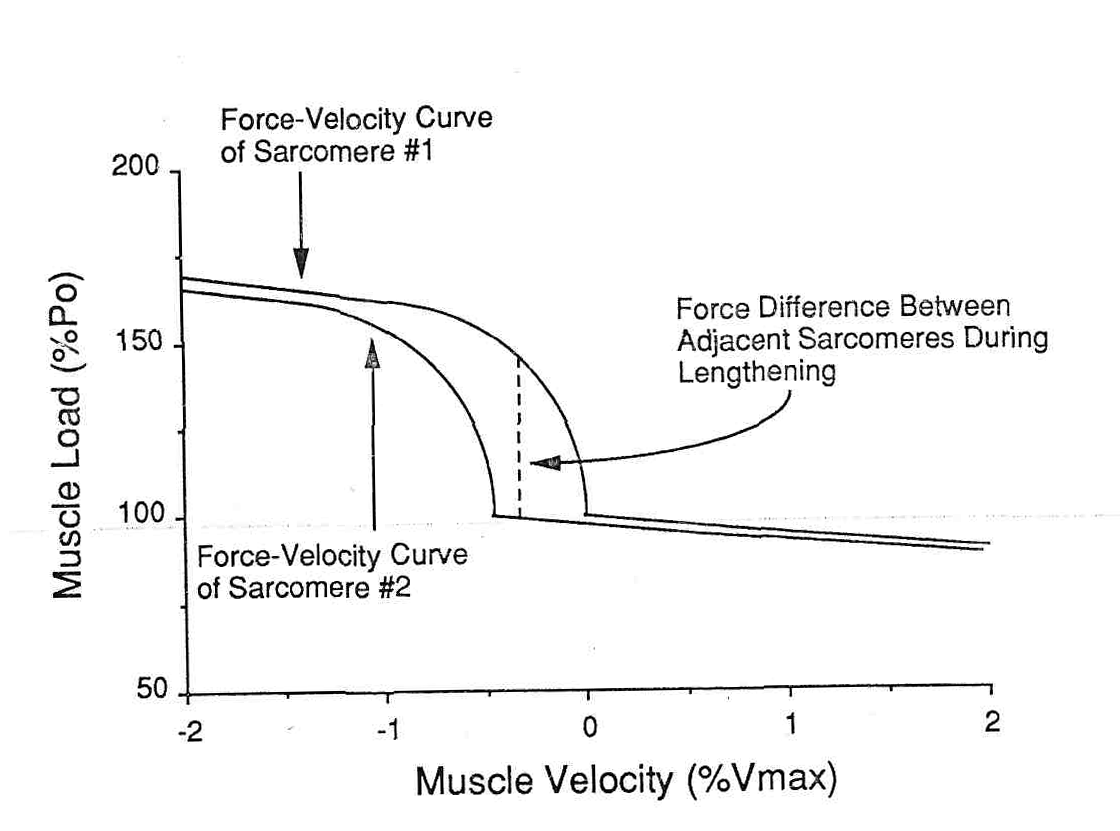
Механическая неустойчивость между смежными саркомерами. Давайте рассмотрим два смежных саркомера, которые имеют немного различные скорости и длину (в пределах физиологического диапазона). Небольшие различия в длине и скорости у укорачивающейся мышцы не несут никакой угрозы, так как отношения кривой «скорость–сила» около нулевой линии не очень круты. Таким образом, для различия Vmax на 0,5 % в течение укорочения, сила, смежных саркомеров изменяется только Pо на 1-2 % (рис. 7). Однако, для смежных саркомеров в течение удлинения, из-за крутизны зависимости «скорость-сила» в смежном саркомере (или даже в смежных областях наложения в пределах саркомера) может измениться больше чем 50%-ым от Pо (рис. 7).
Рис.7. Гипотетические отношения между двумя смежными саркомерами, которые имеют немного различные скорости растяжения. Отметьте, что сила увеличивается очень резко при удлинении и очень небольшие различия в скоростях (0.5%Vmax) могут привести к большим различиям силы между смежным саркомерами. Отметьте, однако, что во время укорочения волокна такого неравенства нет, поскольку отношение между силой и скоростью этой области очень незначительно.
Таким образом, не удивительно, что актиновые филаменты могут испытывать существенно различные силы на своих концах, которые вызовут смещающее напряжение в Z-диске. Возможно, что это смещающее напряжение приводит к «размытию» Z-диска, которое было описано выше. Такой сценарий недавно более подробно рассмотрел Morgan (39), в котором он описывает прогрессивное повреждение смежных саркомеров, поскольку сила волокна превышает их максимальное напряжение растяжения. Если высокое напряжение было бы единственной причиной миофибриллярного разрушения, можно было бы ожидать повреждение, когда пассивное растяжение с высоким напряженным было бы приложено к мышечно-сухожильному соединению. Однако, это, по-видимому, не происходит, так как циклическое пассивное растяжение с напряжением, приближающимся к Pо, изменило максимальное тетаническое напряжение на модели кролика не больше чем 10 % (34). Таким образом, смещение напряжения, а не абсолютное напряжение является более важным в повреждения мышцы. Чтобы проверить гипотезу, что повреждение мышцы непосредственно связано с растяжением мышечного волокна, мы циклически изменяли длину и скорость мышцы tibialis anterior кролика (32). В группе ES (раннее растяжение), мышца был растянута в начале ее активации (рис., величина растяжения составляла 20% от длины волокна, а скорость изменения напряжения составляла 60% в секунду. И величина и степень растяжения были в пределах физиологического диапазона. В группе LS (позднее растяжение), растяжение мышцы было отсрочено на 200 мс, в то время как мышца развивала напряжение. После этого, мышца была растянута так же, как в группе ES. Таким образом, степень растяжения и величина между группами были идентичны. Однако, из-за выбора времени растяжения, пиковая сила, достигнутая в группе LS была почти в два раза больше, чем в группе ES (рис. .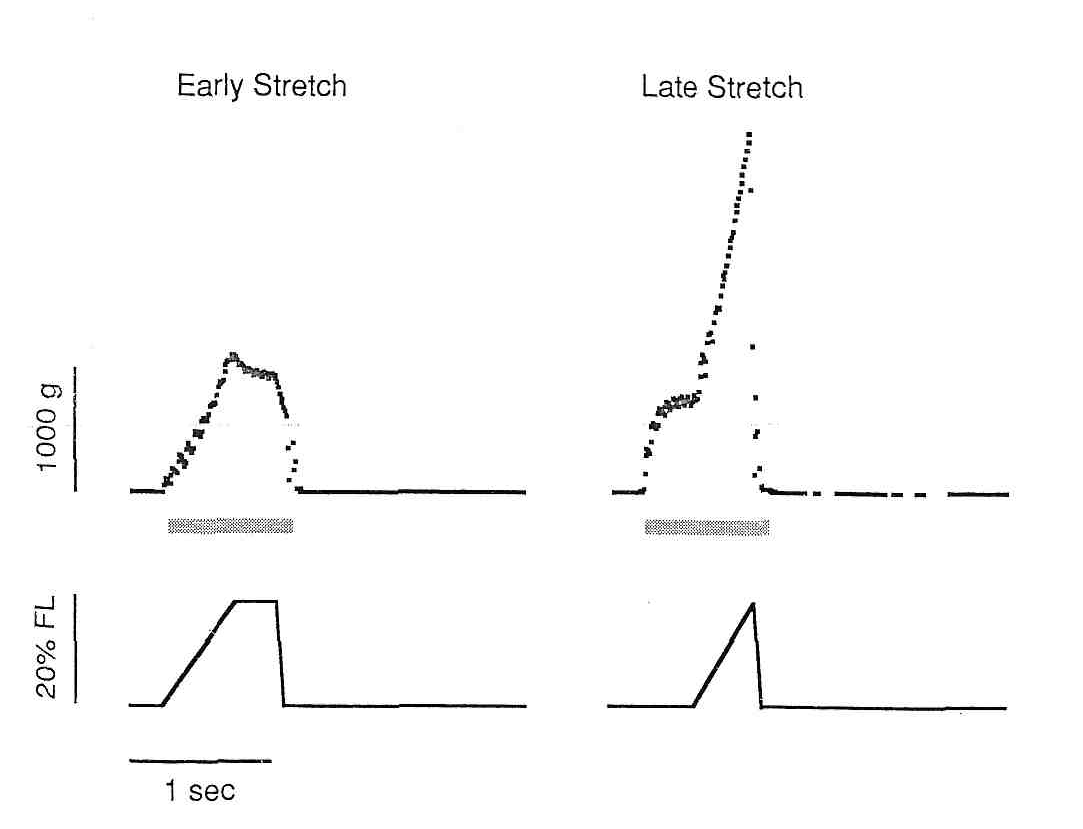
Рис.8. Изменение силы (сверху) и деформации (снизу) в двух экспериментальных группах.Горизонтальная заштрихованная полоса характери зует период в течение готорого мышца стимулировалась
Таким образом, обе группы получили идентичные образцы деформации, но существенно различные напряжения. Интересно, что несмотря на то, что LS группа, дважды стимулировалась для развития полного напряжения в процессе упражнения (30 мин) максимальное тетаническое напряжение после этих двух обработок было почти идентичным (рис. 9). Таким образом, этот эксперимент демонстрирует, что повреждение мышцы, вызванное эксцентричным сокращением, непосредственно не связано с напряжением мышцы. Таким образом, установлено, что напряжение мышцы является высоким в течение эксцентрических сокращений и что повреждение мышцы связано именно с этим типом сокращений, повреждение не обязательно может быть прямой функцией напряжения мышцы. Данные показывают, что саркомер мышцы может иметь физический порог повреждения, который достигается в течение эксцентрического сокращения. В последующем эксперименте, мы варьировали натяжение, приложенное к ТА кролика. У одной группы это натяжение составляло 12.5 % от длины волокна (не от длины мышцы), в то время как у второй группы натяжение составляло 25 % от длины волокна (рис. 9).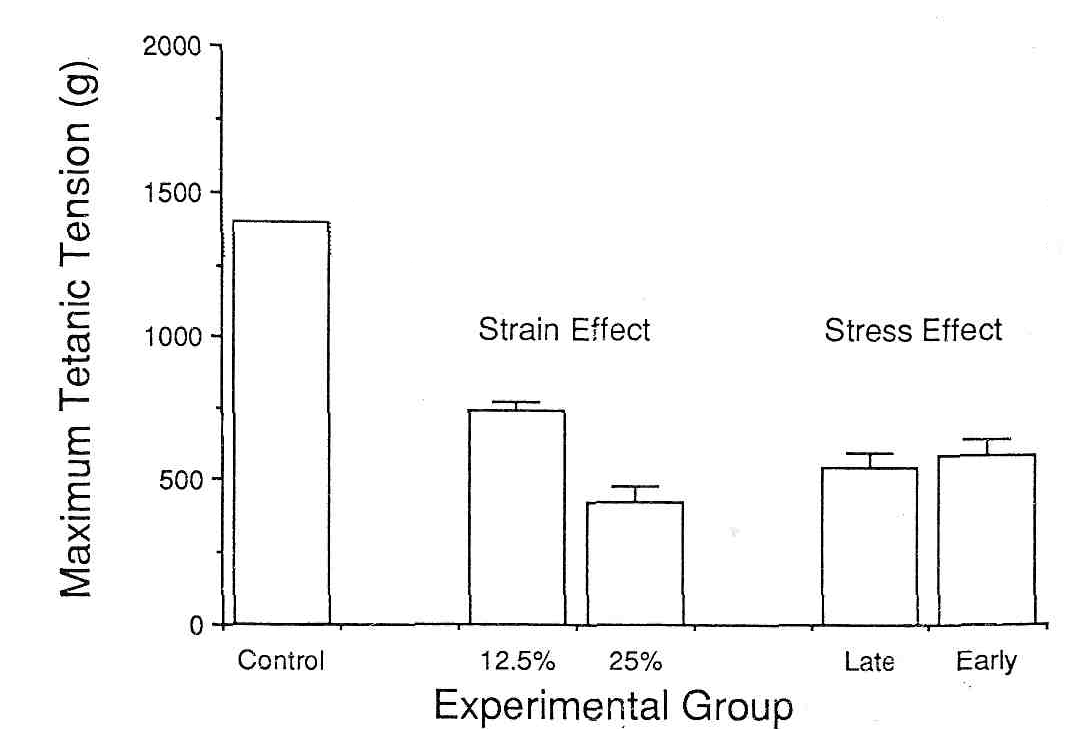
Рис.9 Максимальное тетаническое напряжение tibialis anterior кролика до и после эксцентрических сокращений. Заштрихованная область представляет средние ± SD для нормального напряжения мышцы кролика. Каждый брусок представляет данные для шести — восьми животных.
В отличие от вышеупомянутого эксперимента, изменение натяжения имело совсем другие результаты в терминах мышцы Pо. Маленькое натяжение произвело 45%-ое уменьшение Pо, в то время как увеличенное натяжение произвело 65%-ое уменьшение. Если предположить, что повреждение связано не с напряжением, то нужно принять, что натяжение является важным факторов при повреждении мышцы. Напряжение по существу не является повреждающим фактором, так как так как пассивное растяжение не производит никаких ультраструктурных повреждений и минимальных сократительных изменений (34). Только активное растяжение производит повреждение.Специфика типа волокна и механизм повреждения. В исследовании Friden с соавт. (13), были представлены доказательства, что быстрые волокна повреждаются больше при эксцентрическом сокращении. В том исследовании степень повреждения Z-полосы мышечных волокон IIтипа к I типу было равно 2,8:1 немедленно и 3.0:1 через три дня после упражнения. Мышечные волокна IIВ типа повреждались в 2,5 раза чаще мышечных волокон IIА типа. Нужно, однако, учитывать, что средняя объемная плотность поврежденного материала составляла только 1.6% и 2.4 %, хотя одна треть и половина мышечных волокон были затронуты при выполнении двух биопсий. Данные, свидетельствующие о сильном повреждении быстрых волокон противоречат данным Армстронга с соавт (3), которые при исследовании бега крыс по наклонной плоскости получили данные о сильном повреждении глубоко расположенных волокон первого типа. Однако никакого ультраструктурного исследования типа волокон в том исследовании не было произведено. Это противоречие, возможно связано с несколькими причинами: 1) В исследованиях человека испытуемые могли прекратить упражнение в любое время и в некоторых так и было, когда они ощущали сильный субъективный дискомфорт. Вероятно поэтому в исследованиях на человеке работа в эксцентрическом режиме не приводила к некрозу мышечных волокон. С другой стороны, у животного большой период эксцентрического упражнения конечно вызывал существенные метаболические сдвиги в медленных волокнах 2) в этих исследованиях изучались мышцы с полностью различными функциями. Сопротивление и адаптация к эксцентрическому упражнению может зависеть от различных, к сожалению неизвестных врожденных структурных свойств. 3) возможно, что в глубоком слое, где преобладают медленные мышечные волокна, существуют быстрые мышечные волокна, которые были повреждены. 4) в мышцах большими являются волокна второго типа, а у крыс – волокна первого типа (33). Таким образом, возможно, самые большие волокна наиболее уязвимы для повреждения.
Однако, с ультраструктурной точки зрения, может быть разумно предположить, что более широкие Z-полосы в волокнах I типа менее уязвимы при повторных, высоко интенсивных упражнениях, чем волокна II типа. В морфологическом исследовании бегунов, выполняющих анаэробную работу на тредмиле, мы выполнили полное ультраструктурное изучение поврежденных волокон (11). В этом исследовании была осуществлена специальная фотографическая техника. Эта процедура состояла из проектирования увеличенного изображения неповрежденной части поврежденного волокна через апертуру разреза. Эта апертура после этого перемещалась вдоль фотографической бумаге с постоянной скоростью. Направление движения было параллельно поперечным линиям миофибрилл. Используя эту технику, число и плотность М мостов так же, как ширина Z-полосы могли ясно визуализироваться и использоваться как критерии для ультраструктурной классификации волокон. Через два часа после повторных попыток спринтерского бега в 36% волокон m.vastus lateralis были обнаружены повреждения. Самые заметные изменения были снова связаны с Z-дисковыми отклонениями с центральным и обширным вытеканием. Из поврежденных волокон 80% отклонений были найдены в мышечных волокнах с узкими Z-полосами и имеющих только три М полоски, то есть в волокнах IIB типа. Интересно, биопсия тех же волокон и последующее исследование тех же самых волокон показали истощение гликогена в мышечных волокна I типа и IIB типа, но не в волокнах IIA типа. Однако только в 20 % общего количества мышечных волокон было обнаружено полное истощение гликогена. Это наблюдение может указать существенное метаболическое требование к волокнам IIB типа, которое, в дополнение к эксцентрической нагрузке при беге на длинную дистанцию, увеличивает риск повреждения более тонкого, и возможно более слабого Z-диска волокон IIB типа.
В связи с тем, что недавние исследования на животных показали, что все волокна мышцы, которые имели небольшие повреждения были волокнами FG типа, мы подумали, что окислительная способность мышц могла бы быть причастна к первичным повреждениям мышцы (34). (Отметьте, что метаболическая схема классификации, используемая для мышцы животных – аналогична, но не идентичная базирующейся на активности АТФ-азы схеме классификации мышечных волокон человека). Таким образом, типы SO, FOG, FG аналогичны МВ I типа, IIA типа, IIB типа. Если окислительная способность волокна мышцы, является одним из факторов повреждения волокна, мы могли бы выдвинуть гипотезу о схеме повреждения, которая происходит следующим образом: в ранний период упражнения (в пределах первых 10 минут), наблюдается утомление волокон типа FG. Из-за их неспособности к ресинтезу АТФ, у них наблюдается ригор или высокая жесткость. Последующее растяжение жестких волокон приводит к наблюдаемому разрушению цитоскелета и повреждению миофибрилл. Эта гипотеза поддерживается рядом фактов. Во-первых, она объясняет хорошо известный "защитный" эффект, который связан с тем, что тренировка на выносливость предотвращает повреждения вызванные эксцентрической работой. Тренировка на выносливость, как известно приводит к увеличению окислительной способности мышц и поэтому, волокна типа FG превращаются в волокна типа FOG. Поскольку волокна FOG типа не утомляются и входят в ригор, также, как и волокна FG типа, тренировка на выносливость приводит к меньшим повреждениям, связанным с эксцентрическим сокращением мышц. Понимание этого может быть важным в тренировочных режимах. Эту гипотезу можно экспериментально проверить. Мы предсказываем, что повреждение, вызванное эксцентрическим режимом сокращения мышцы, происходит в первые минуты тренировки. Мы также предсказываем, что утомленная мышца (например, после 20 минут изометрического сокращения) могла бы быть повреждена более значительно, если бы эксцентрическое растяжение было предпринято в конце тренировочного периода. Повреждение не обязательно требовало бы числа растяжений. Теперь остается проверить эти гипотезы.
Второй механизм повреждения, который мог зависеть от окислительной способности волокна, имеет отношение с другими клеточными процессами, которые основаны на окислительном метаболизме. Например, большое уменьшение adenylate мышечного волокна может уменьшить митохондриальную способность противостоять ионам кальция, приводя к активации активизированных кальцием нейтральных протеаз. Интересно, что эти ферменты были недавно вовлечены в процесс дегенерации – регенерации мышечного волокна (4).
РЕЗЮМЕ
Повреждение мышцы, вызванное эксцентрическим сокращением, наблюдается на животных и людях. Хорошо видны повреждения цитоскелета и миофибрилл. Хотя повреждения, связанные с эксцентрическими повреждениями, связаны также с высоким напряжением, возможно большое напряжение само по себе не вызывает повреждений. Активное растяжение, кажется, лучше коррелирует с повреждением. Больше повреждаются быстрые мышечные волокна, имеющие низкую окислительную способность. Это дает возможность предположить, что окислительная способность – важный фактор в повреждении мышцы, вызванном эксцентрическими сокращениями.
#10761
Отправлено 30 ноября 2012 - 12:53
roma2
-
- Привилегированный пользователь
-
- 9 032 сообщений
Чатланин Уэф
-
Имя: Уэф -
Пол: Мужчина -
Город: г. "Трех шурупов", Планета Плюк, 215 в Тентуре, галактика Кин-дза-дза в Спирали
#10762
Отправлено 30 ноября 2012 - 01:06
Deejgrey
-
- Участник форума
-
- 182 сообщений
Новичок
-
Имя: Сергей -
Пол: Мужчина -
Город: Евпатория
#10763
Отправлено 30 ноября 2012 - 01:10
sandwolf77
-
- Модератор
-
- 21 009 сообщений
crusader of philosophy
-
Имя: Алексей -
Пол: Мужчина -
Город: Тюмень
А.Самсонова у себя в работе считает, что причина повреждаемости волокон 2б типа в более тонких z-дисках. но какая,в принципе, разница, что в лоб, что по лбу, так и так травмируются первыми. кстати, в работе много опытов по повреждаемости волокон, но что-то не заметил опытов по доказательству, что именно разрушения(которые теперь уже вроде факт), лучший стимул роста. ведь интересно, механосигналинг - это очень малое травмирование, которое накапливается либо перемещается в разные мыш.волокна(это я образно, если что), или реально без повреждений сам натяг срабатывает?Это дает возможность предположить, что окислительная способность – важный фактор в повреждении мышцы, вызванном эксцентрическими сокращениями.
Сообщение изменено: sandwolf77 (30 ноября 2012 - 01:11)
#10764
Отправлено 30 ноября 2012 - 01:13
roma2
-
- Привилегированный пользователь
-
- 9 032 сообщений
Чатланин Уэф
-
Имя: Уэф -
Пол: Мужчина -
Город: г. "Трех шурупов", Планета Плюк, 215 в Тентуре, галактика Кин-дза-дза в Спирали
В лабораторных условиях издеваться над мышками эксцентрикой не считаем!Но ссылки касаемо животных у автора имеются.
#10765
Отправлено 30 ноября 2012 - 08:17
Еретик
-
- Участник форума
-
- 1 125 сообщений
Приведи, хоть один пример эксцентрического сокращения в животном мире!
Или уж сразу негативного оверлоада!
А мона я приведу?
Приземление после прыжка, тем более, с большой высоты. Особенно впечатляют прыжки крупных древолазов вроде леопарда.
Торможение после быстрого бега.
Притормаживание крупных зверей при быстром спуске по склону. Например, мудведь вверх по склону бежит быстрее, чем вниз - туша большая, центр тяжести при спуске падает на относительно слабые передние лапы.
Удержание крупной добычи, по неизвестным причинам не желающей быть съеденной.
Вырывание еды из пасти/лап собрата.

Сообщение изменено: Еретик (30 ноября 2012 - 08:27)
#10766
Отправлено 01 декабря 2012 - 08:14
Михалы4
-
- Участник форума
-
- 25 967 сообщений
body_by_mma
-
Имя: Антон -
Пол: Мужчина -
Город: Москва
некоаксиальность миофибрилл
Коаксиальность – по-русски означает "соосность", т.е. расположение осей на одной линии.
Вопрос: разве в отношении миофибрилл можно говорить о какой-то некоаксиальности?
З.Ы. Читаем далее...
Сообщение изменено: Михалы4 (01 декабря 2012 - 08:16)
#10767
Отправлено 04 декабря 2012 - 07:10
Disco
-

- Привилегированный пользователь
-
- 10 510 сообщений
ПериодизаторЪ
-
Имя: Александр -
Пол: Мужчина -
Город: Ривендейл
100-end
НЕ ФЛУДИТЕ!
"Думайте обо всём , что пишете ... , а не пишите все, о чём думаете!"

Все посты Priest.72, ARB, KomRADik, Михалы4, и остальных "активистов-дискуссантов" отредактированы путем объединения 2-х и более постов в один, по причине их близости в теме после зачистки топика от флуда. Содержимое постов оставлено без изменения за исключением редких случаев (мат, спряжения, падежи, приведение части поста в соответствие с логической линией предыдущего автора).
P.S. Андрей Юрьевич! ПИШИТЕ ПОСТЫ СРАЗУ, а не по 5 в ряд ... как дроп! 
Сообщение изменено: Disco (04 декабря 2012 - 12:10)
#10768
Отправлено 07 декабря 2012 - 12:01
ctjan
-
- Участник форума
-
- 6 453 сообщений
Бывалый
-
Пол: Не определился -
Город: где то рядом
The combination of creatine monohydrate supplementation (CR) and resistance
training has been shown to synergistically accentuate muscle fiber hypertrophy [1,2]
and muscle cross-sectional area (CSA) [1]. Several studies demonstrated that CR
supplementation was effective for increasing lean muscle mass, strength, muscular
power, and hydration status [3-7]. Kilduff et al. [8] demonstrated that four weeks of
CR supplementation in conjunction with resistance training increased maximal
strength more than resistance training alone. Jonhson et al. [9] examined the influence
of a loading phase of CR (20g/day for 6 days) on bilateral leg extension repetition
performance (concentric and eccentric muscle actions) until voluntary exhaustion in
18 men and women. The results indicated an approximate increase of 25% and 15%
from baseline for the dominant leg in men and women, respectively. From a
longitudinal standpoint, Huso et al. [10] demonstrated that 12 weeks of CR
supplementation combined with resistance training increased body mass and muscle
mass more than resistance training alone.
"Strength and hypertrophy responses to constant and decreasing rest intervals
in trained men using creatine supplementation Journal of the International Society of Sports Nutrition 2011"
кто там про воду говорил)
training has been shown to synergistically accentuate muscle fiber hypertrophy [1,2]
and muscle cross-sectional area (CSA) [1]. Several studies demonstrated that CR
supplementation was effective for increasing lean muscle mass, strength, muscular
power, and hydration status [3-7]. Kilduff et al. [8] demonstrated that four weeks of
CR supplementation in conjunction with resistance training increased maximal
strength more than resistance training alone. Jonhson et al. [9] examined the influence
of a loading phase of CR (20g/day for 6 days) on bilateral leg extension repetition
performance (concentric and eccentric muscle actions) until voluntary exhaustion in
18 men and women. The results indicated an approximate increase of 25% and 15%
from baseline for the dominant leg in men and women, respectively. From a
longitudinal standpoint, Huso et al. [10] demonstrated that 12 weeks of CR
supplementation combined with resistance training increased body mass and muscle
mass more than resistance training alone.
"Strength and hypertrophy responses to constant and decreasing rest intervals
in trained men using creatine supplementation Journal of the International Society of Sports Nutrition 2011"
кто там про воду говорил)
#10769
Отправлено 08 декабря 2012 - 01:16
MikeKazakov
-

- Официальное лицо
-
- 22 236 сообщений
МС СССР
-
Имя: Mike -
Пол: Мужчина -
Город: Jerusalem
А где написано, что НЕТ. Приём креатина в СОЧЕТАНИИ С ТРЕНИРОВКАМИ дали прибавку в силе и в массе больше, чем без, ничего нового и интересного. С 20гр креатина в день можно почки в унитаз спустить, если что. Где там прирост в силе и массе на креатине БЕЗ тренировок, как с гормонами?кто там про воду говорил)
#10770
Отправлено 08 декабря 2012 - 09:11
ctjan
-
- Участник форума
-
- 6 453 сообщений
Бывалый
-
Пол: Не определился -
Город: где то рядом
А где написано, что НЕТ. Приём креатина в СОЧЕТАНИИ С ТРЕНИРОВКАМИ дали прибавку в силе и в массе больше, чем без, ничего нового и интересного. С 20гр креатина в день можно почки в унитаз спустить, если что. Где там прирост в силе и массе на креатине БЕЗ тренировок, как с гормонами?
Есть факт. Интерпретаций можно сказать много. Но мне не известно фактов например что выростит ММ от приема гуараны с тренировками.
Под постом было написано к чему это обращено. Если бы все было известно тогда бы таких ответов не было бы.
2 посетителей читают эту тему
0 пользователей, 2 гостей, 0 скрытых


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}