Вход
Вход Регистрация
Регистрация





Наоборот жеж. Сначала жали, потом ТВБ. В первом сете ТВБ пропал повтор. Во втором случае ТВБ шел сразу за бицепсом. Все тот же пропавший повтор.
Без разницы. На перехлёст я тебе указал.


 74 голосов
74 голосов

Невыступающий неспортсмен


Наоборот жеж. Сначала жали, потом ТВБ. В первом сете ТВБ пропал повтор. Во втором случае ТВБ шел сразу за бицепсом. Все тот же пропавший повтор.
Без разницы. На перехлёст я тебе указал.
IgG к SARS-CoV-2

свежак от Кригера по тестостерону и гипертрофии
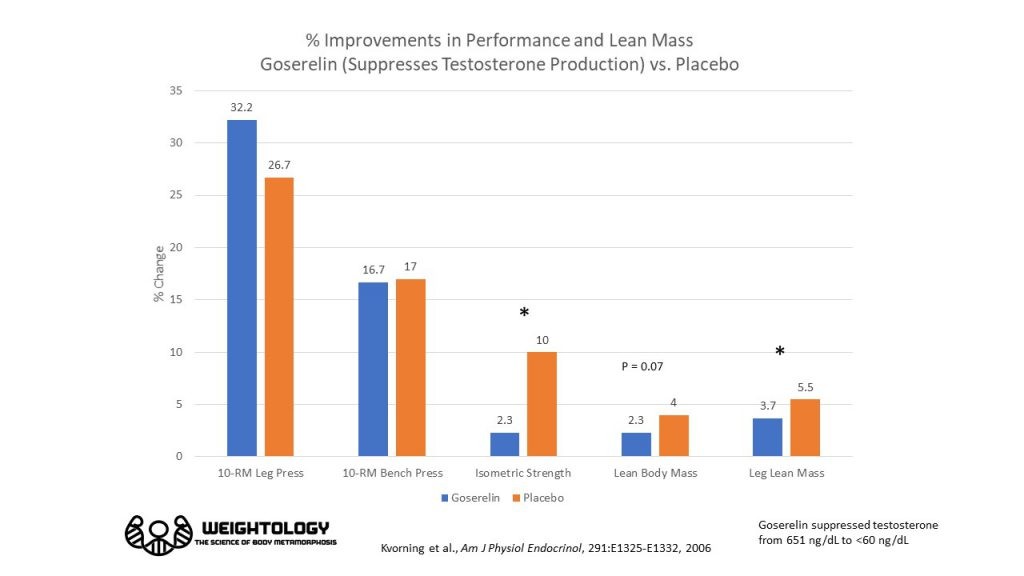
Прикол. Кастраты увеличили жим ногами больше чем плацебники. В жиме лежа не отличались.
Аксакал

свежак от Кригера по тестостерону и гипертрофии
Вроде Анатолий это постил уже выше.
IgG к SARS-CoV-2
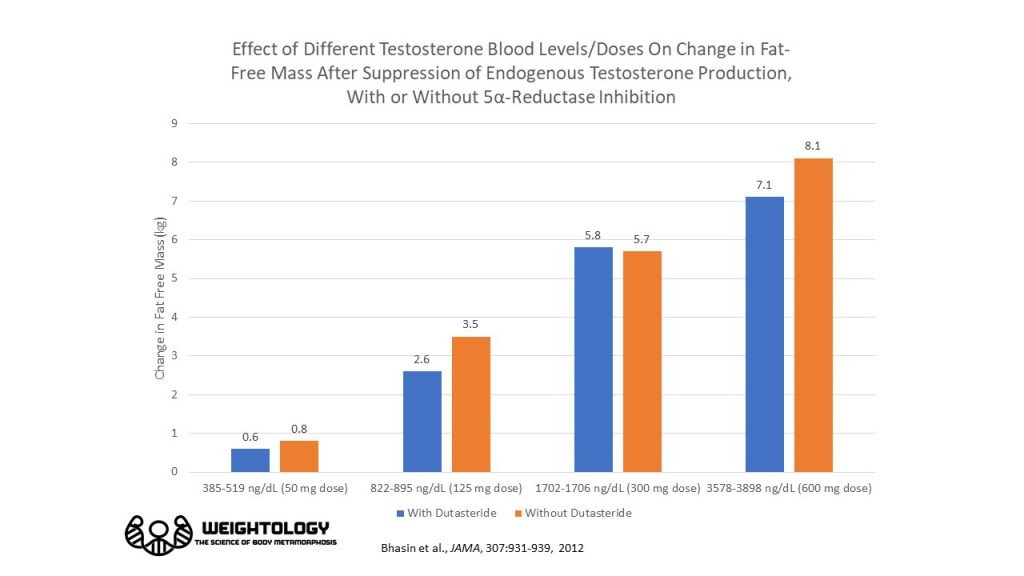
Собирая все это вместе, данные показывают, что масса без жира будет увеличиваться на 0,7 - 1,3 фунта (0,3 - 0,6 кг) на каждые 100 нг / дл повышения уровня тестостерона в крови в пределах физиологического диапазона. Таким образом, если вы перешли с 300 нг / дл до 600 нг / дл, это будет 0,9–1,8 кг или примерно 2,1–4 фунта.
Грубо говоря при разгоне 4->8 (вдвое!) получим халявных 2 кг мышц. ВСЕГО ЛИШЬ!
Так что Рихаду стоит замолчать навсегда про кломид.
IgG к SARS-CoV-2
Кстати у этого типа есть интересные новые исследования
https://www.science....le/phillis.html
IgG к SARS-CoV-2
The impact of exercise and nutrition in the regulation of skeletal muscle mass. - это вообще мегатруд.
Книжка цельная даже есть
https://www.elsevier...8-0-12-810422-4
Сообщение изменено: oscid (14 августа 2020 - 06:37)
IgG к SARS-CoV-2
Регулирование массы скелетных мышц с помощью упражнений и кормления
Изучение биологических механизмов, регулирующих размер мышечной массы человека, было и остается предметом интенсивных научных исследований. Более 30 лет назад профессор Майк Ренни и его коллеги (Ренни и др., 1982) впервые продемонстрировали, что скорость включения внутривенно введенного стабильного меченого изотопом аминокислотного индикатора в белок скелетных мышц стимулируется кормлением. По сравнению с ночным голоданием, скорость синтеза мышечного протеина (MPS) удваивалась после приема пищи, а увеличение MPS после приема пищи составляло большую часть положительного чистого белкового баланса на уровне всего тела (Rennie et al., 1982). С тех пор в серии исследований, проведенных группой профессора Ренни (например, Rennie et al. 2004; Atherton et al. 2010) и других (например, Biolo et al. 1995, 1997; Reidy et al. 2017), изучалась суточная регуляция мышечной ткани. белковый оборот (например, баланс между синтезом и распадом). Из этой работы мы узнали, что во время базальных, постабсорбтивных состояний скорость распада мышечного белка (MPB) превышает скорость MPS, вызывая чистую потерю белка (Rennie et al. 2004), и потребление пищи компенсирует эти потери, потому что диета Аминокислоты, полученные из белков, стимулируют MPS, а инсулин подавляет MPB (Rennie et al. 2004). Чистый прирост белка после приема пищи в значительной степени определяется количеством потребляемого белка, потому что результирующее увеличение незаменимых аминокислот в плазме стимулирует MPS дозозависимым образом до ~ 30 г пищевого белка (или эквивалентного количества аминокислот) (Cuthbertson et al. 2005; Moore et al. 2015), тогда как концентрация инсулина, необходимая для достижения максимального подавления MPB (∼15–30 мЕд / л), уже возникает после употребления небольшого количества белка или углеводов (Bohe et al. 2001; Гринхафф и др. 2008; Мур и др. 2009). Соответственно, простое употребление большего количества белка приводит к увеличению скорости окисления аминокислот без дальнейшего увеличения MPS (Moore et al. 2015).
Помимо белково-аминокислотного питания, упражнения служат другим основным анаболическим стимулом для скелетных мышц. Классическая работа Ренни и др. (1981) показали, что интенсивная тренировка приводит к улучшению баланса чистого белка всего тела после тренировки за счет увеличения синтеза белка в организме, превышающего его распад. Спустя более десяти лет аналогичные результаты были сделаны на уровне скелетных мышц, показывающие, что один сеанс упражнений с отягощениями увеличивал как MPS, так и MPB на срок до 48 часов, но хотя относительная стимуляция MPS была больше, чем MPB, MPB все же превышала MPS натощак, не приводящий к приросту чистого мышечного белка (Phillips et al. 1997). Важно отметить, что эффекты упражнений и приема протеина складываются (Witard et al. 2009; Pennings et al. 2011). Таким образом, физические упражнения увеличивают мышечную массу в основном из-за увеличения MPS, а не подавления MPB.
Помимо белково-аминокислотного питания, упражнения служат другим основным анаболическим стимулом для скелетных мышц. Классическая работа Ренни и др. (1981) показали, что интенсивная тренировка приводит к улучшению баланса чистого белка всего тела после тренировки за счет увеличения синтеза белка в организме, превышающего его распад. Спустя более десяти лет аналогичные результаты были сделаны на уровне скелетных мышц, показывающие, что один сеанс упражнений с отягощениями увеличивал как MPS, так и MPB на срок до 48 часов, но хотя относительная стимуляция MPS была больше, чем MPB, MPB все же превышала MPS натощак, не приводящий к приросту чистого мышечного белка (Phillips et al. 1997). Важно отметить, что эффекты упражнений и приема протеина складываются (Witard et al. 2009; Pennings et al. 2011). Таким образом, физические упражнения увеличивают мышечную массу в основном из-за увеличения MPS, а не подавления MPB.
Старение приводит к характерным изменениям в обмене мышечного белка, называемым возрастной «анаболической резистентностью» (Moore et al. 2015), которая характеризуется притупленной реакцией как MPS, так и MPB на анаболические эффекты аминокислот и упражнений и антипротеолитический эффект инсулина, приводящий к постепенной потере мышечной массы. Первое свидетельство этого феномена было предоставлено профессором Ренни и его коллегами (Катбертсон и др., 2005; Уилкс и др., 2009; Кумар и др., 2009a), которые продемонстрировали, что, хотя базальные скорости МПС и МПБ не различаются у молодых и у пожилых людей увеличение MPS в результате приема аминокислот уменьшается у пожилых людей. Позже они продемонстрировали, что опосредованное инсулином подавление MPB (Wilkes et al. 2009) и вызванное физическими упражнениями увеличение MPS (Kumar et al. 2009b) также снижаются у пожилых людей по сравнению с молодыми людьми. Наличие анаболической резистентности у пожилых людей с тех пор подтверждено несколькими другими исследователями (Moore et al. 2015). Кроме того, эта анаболическая резистентность больше поражает пожилых женщин, чем мужчин (Smith et al. 2012). Поэтому стратегии профилактики и лечения возрастной саркопении сосредоточены на преодолении этой анаболической резистентности (Bauer et al. 2013). Эти усилия привели к новым открытиям, связанным с потенциальной важностью небелковых пищевых компонентов для регуляции обмена мышечного белка.
Ремоделирование мышечного белка после употребления богатой белком пищи
Хотя мы многое узнали о регуляции обмена мышечного белка из исследований, в которых вводили изолированный белок или аминокислоты, пища - это то, что люди едят, и стало очевидно, что, хотя белок / аминокислоты являются основным анаболическим стимулом для мышц, источником диетический белок и взаимодействие с пищевым матриксом могут иметь важное влияние на биологическую активность белка / аминокислот. В то время как потребление богатого белком постного мяса, как было обнаружено, стимулирует MPS зависимым от дозы криволинейным образом как у молодых, так и у пожилых людей (Symons et al. 2009; Robinson et al. 2013), а реакция MPS на употребление нежирной говядины была максимальной. с размером порции ∼113–170 г (4–6 унций), что обеспечивает ∼30–35 г белка. Исследования белков цельных продуктов редки, но проводились. Например, употребление изонитрогенного обезжиренного молока по сравнению с нежирной (97% обезжиренной) говядиной привело к большей доступности циркулирующих пищевых аминокислот после употребления говядины, чем молока в ранней постпрандиальной фазе (0–2 часа), но, как это ни парадоксально, MPS Ответ был больше после молока, чем после говядины в это время (Burd et al. 2015). При оценке в течение 5-часового периода не наблюдалось различий в показателях MPS между постной говядиной и обезжиренным молоком, несмотря на тенденцию к более высокой доступности аминокислот в рационе после употребления говядины. Недавнее сравнение потребления яичного белка и цельного яйца (оба обеспечивают 18 г пищевого белка) во время восстановления после тренировки у здоровых молодых мужчин показало превосходную 5-часовую реакцию MPS после приема цельного яйца, несмотря на аналогичный постпрандиальный аминокислотный профиль после приема внутрь эти два источника белка (van Vliet et al., 2017). Эти результаты подтвердили результаты предыдущего исследования, в котором потребление цельного молока привело к большему чистому поглощению аминокислот скелетными мышцами (предположительно для использования в синтезе белка) по сравнению с потреблением обезжиренного молока, соответствующего содержанию белка (Elliot et al. 2006 ).
Помимо аминокислот, продукты, богатые белком, часто также содержат широкий спектр липидов, витаминов, минералов и других биоактивных соединений (факторы роста, пептиды, миРНК и т. Д.; Moller et al. 2008; Zanovec et al.2010; Phillips et al. 2015), которые, по-видимому, способны модулировать реакцию MPS при приеме внутрь. Например, на животных моделях было продемонстрировано, что поступление олеата (Tardif et al., 2011), витаминов A, D и E, а также минералов селен и цинк также может напрямую влиять на анаболический ответ мышц (Zhao et al. 2016; Chanet et al.2017). Более того, Smith et al. продемонстрировали, что 8 недель приема добавок омега-3 жирных кислот, полученных из рыбьего жира, увеличивают содержание EPA и DHA в фосфолипидах скелетных мышц и повышают MPS в условиях гипераминоацидемии-гиперинсулинемии как у пожилых (Smith et al. 2011a), так и у молодых (Smith et al. 2011b). Те же авторы расширили эту работу, продемонстрировав, что добавление жирных кислот омега-3, полученных из рыбьего жира, увеличивает массу и функцию скелетных мышц у здоровых пожилых людей (Smith et al. 2015). Этот отчет подтвердил выводы других исследователей, которые показали, что диетические добавки с омега-3 жирными кислотами способствовали увеличению силы скелетных мышц во время тренировок у пожилых женщин (Rodacki et al. 2012; Da Boit et al. 2017). Биологические механизмы, с помощью которых эти жирные кислоты могут регулировать анаболизм скелетных мышц, остаются неизвестными. Включение в рацион высоконенасыщенных жирных кислот EPA и DHA было связано с усилением механистической мишени передачи сигналов комплекса-1 рапамицина (Smith et al. 2011a; Kamolrat et al. 2013; McGlory et al. 2014), что дает один потенциальный шанс механизм, с помощью которого можно увеличить анаболизм. Также возможно, что включение омега-3 жирных кислот в скелетные мышцы изменяет формирование липидного растра (Hou et al., 2016) и передачу механических и питательных сигналов трансляционному аппарату. Таким образом, жирные кислоты омега-3 представляют собой многообещающую область исследований для разработки стратегий питания для противодействия анаболической резистентности у пожилых людей. Следующим логическим шагом будет исследование того, может ли употребление жирных кислот, таких как омега-3, компенсировать атрофию скелетных мышц в периоды постельного режима / неиспользования мышц у пожилых людей. Такие данные неизбежно будут иметь важное клиническое значение, учитывая распространенные метаболические последствия периодического неиспользования мышц у пожилых людей (English & Paddon-Jones, 2010; McGlory et al., 2017).
Таким образом, мы только недавно начали понимать и, возможно, ценить сложность различных источников пищи и их роль в стимулировании ремоделирования протеома мышц. Очевидно, что требуется гораздо больше работы, чтобы понять, как прием пищи, в отличие от одного только белка / аминокислот, может повлиять на суточную потребность в белке, особенно у пожилых людей. Однако имеющиеся данные позволяют предположить, что наличие витаминов, минералов, липидов и других биоактивных питательных веществ в пище работает вместе с аминокислотами / белками, поддерживая постпрандиальный рост MPS. Выяснение активных биологических компонентов небелковых продуктов питания, такие как видовые липидных, может дать важные данные, имеющие отношение к разработке новых методов лечения нутрицевтиков способствовать костно-мышечной системе у пожилых людей.
Протеомный / фосфопротеомный анализ обмена мышечного белка
Традиционные методы оценки белкового обмена у людей основаны на инфузии стабильных изотопно-меченых аминокислот в сочетании с серийными биопсиями мышц. Образцы мышечной биопсии анализируются на предмет включения индикатора в смешанные или специфические фракции мышечного белка (например, миофибриллярные, саркоплазматические, митохондриальные) или несколько отдельных белков (например, актин или тяжелая цепь миозина). До недавнего времени оставалось неизвестным, как упражнения с отягощениями и питание влияют на скорость обмена многочисленных отдельных белков, составляющих скелетные мышцы. Пытаясь восполнить этот пробел в знаниях, Мерфи и его коллеги (2018) вводили дейтерированную воду (D2O) своим участникам исследования и использовали тандемно-масс-спектрометрический протеомный анализ образцов мышечной биопсии (Шанкаран и др., 2016b), чтобы выявить изменения в синтетической воде. уровни> 150 отдельных мышечных белков, содержащихся во фракциях миофибриллярных, митохондриальных и саркоплазматических белков. Первичный результат этого исследования (Мерфи и др., 2018) заключался в том, что пожилые люди, выполняющие упражнения с отягощениями в состоянии ограниченного потребления энергии, усиливали синтез 175 из 195 измеренных индивидуальных белков скелетных мышц в каждой из конкретных фракций белка. Помимо активации отдельных белков скелетных мышц в миофибриллярной (т. Е. Сократительной) категории, был также синтез белков, участвующих в регуляции многочисленных метаболических процессов, таких как гликолиз (т. Е. 6-фосфофруктокиназа) и цепь переноса электронов (т. Е. Субъединицы АТФ-синтазы). увеличивается после упражнений с отягощениями.
Примерно в то же время, когда Мерфи и его коллеги (2018) сообщили о своих результатах, Камер и коллеги (2017a) применили метод динамического профилирования протеома для оценки влияния упражнений с отягощениями во время диеты с высоким содержанием жиров и низким содержанием углеводов на синтез и распад отдельные мышечные белки у молодых людей. Они обнаружили, что изобилие 28 из 90 измеренных миофибриллярных и саркоплазматических белков увеличилось, и что это было связано как с усилением синтеза, так и со снижением скорости распада этих белков. Интересно, что первоначальное увеличение содержания белка после упражнений с отягощениями произошло практически без изменений в синтезе белка, что подчеркивает потенциальную ключевую роль MPB в ремоделировании протеома мышц при упражнениях с отягощениями. Такие результаты могут противоречить существующим знаниям о том, что изменения протеома при упражнениях с отягощениями в основном вызваны изменениями в скорости MPS. Однако важно интерпретировать эти результаты с некоторой осторожностью, так как скорость распада оценивалась по разнице между изменением количества белка и его синтезом с течением времени. Таким образом, распад белка не оценивался напрямую, и 11 белков даже продемонстрировали физиологически невозможное отрицательное значение для расчетной скорости распада, что подчеркивает необходимость дальнейшего совершенствования подходов `` омикс '', используемых для изучения роли MPB в адаптивной реакции скелетных мышц на физические нагрузки. активность и диетические вмешательства. Как признают авторы, их выводы, касающиеся MPB, могут быть связаны с техническими ошибками, но также возможно, что рециркуляция аминокислот могла увеличивать изобилие белка в большей степени, чем предсказал синтез. Тем не менее, это исследование (камера и др. 2017a) демонстрирует мощь динамического протеома профилирования применительно к исследованиям скелетных мышц человека, который может быть использован, чтобы обнаружить новые и клинический важные мероприятия по улучшению здоровья опорно-двигательного аппарата. Например, одним из выводов исследования (Camera et al., 2017b) было то, что в ответ на упражнения с отягощениями наблюдалось значительное увеличение синтеза белка тяжелой цепи миозина, но только синтеза быстро сокращающегося белка 2a (MYHC2), который является захватывающее открытие, учитывая, что саркопения характеризуется снижением экспрессии быстро сокращающихся волокон (Lexell et al. 1988).
Хотя достижения в области молекулярной / клеточной биологии за последние 20 лет позволили получить важную информацию о регуляции обмена мышечного белка, наше понимание механизмов, ответственных за потерю мышечной массы с возрастом, остается неуловимым. Имеются доказательства того, что неспособность адекватно активировать mTORC-1 и впоследствии транслировать гены, кодирующие мышечные белки, играет некоторую роль. Однако такие сигнальные сети многогранны и взаимосвязаны, и раскрытие сложности этих взаимодействий вряд ли может быть достигнуто за счет использования только современных подходов, таких как иммуноблоттинг или иммуногистохимия. Одно интересное направление исследований, которое может дать некоторые ответы, - это фосфопротеомика. Фосфопротеомика обеспечивает глобальный и беспристрастный подход к изучению изменений в сетях фосфорилирования в ответ на анаболическую стимуляцию. Например, в двух недавних исследованиях после тренировки на выносливость были выявлены новые сайты фосфорилирования на многочисленных протеинкиназах.
Будущие направления
Со времени первых исследований белкового обмена, которые основывались на подходе артерио-венозного баланса и измерении включения меченых аминокислот в мышечные белки, были достигнуты значительные успехи в нашем понимании факторов, которые регулируют обмен мышечного белка после упражнений и питания. Следующая эра научного прогресса в этой области, несомненно, будет подкреплена использованием «атомных» технологий (например, липидомных, протеомных, метаболиомных, транскриптомных). Основное преимущество применения «атомных» технологий для изучения оборота мышечного белка и связанных внутриклеточных сигнальных сетей состоит в том, что влияние стратегии питания и упражнений можно изучить более подробно и в более широком масштабе. Беспристрастный глобальный анализ изменений кинетики мышечного белка и молекулярных факторов, регулирующих эти изменения, поможет точно определить возрастные и связанные с заболеванием изменения во всех биологических сетях. В настоящее время также есть доказательства того, что белки, собранные в мышцах, такие как креатинкиназа M-типа и карбоангидраза, уходят в кровоток, и что скорость фракционного оборота этих белков в плазме может использоваться в качестве суррогатного маркера обмена мышечного белка (Шанкаран и др., 2016a; Мерфи и др., 2018). Такие данные открывают захватывающую возможность изучения влияния физических упражнений и питания на обмен мышечного белка с помощью минимально инвазивных методов (забор крови). Это особенно важно при работе с больными людьми, например, в отделениях интенсивной терапии или в других столь же опасных клинических ситуациях, для которых взятие биопсии скелетных мышц нецелесообразно или невозможно. Еще одна область, заслуживающая дальнейшего изучения, - это изучение того, как изменения в составе липидов, специфичных для мышц, влияют на обмен мышечного белка. Открытие того факта, что омега-3 жирные кислоты делают скелетные мышцы более «анаболически чувствительными», было первым шагом в этом направлении и требует исследования механизмов, ответственных за их благотворное влияние на обмен мышечного белка, что может включать улучшение текучести мембран и / или изменения в формирование липидного рафта. Дальнейшая работа по управлению содержанием и составом видов фосфолипидов как на доклинических, так и на человеческих моделях, несомненно, поможет дать ответы.
В заключение скажу, что протеом стареющих мышц - это область интенсивных научных исследований. Понимая влияние физической активности и различных источников питательных веществ на метаболизм мышечного белка человека на индивидуальном уровне белка, можно будет разработать более эффективные стратегии противодействия нормальной возрастной потере мышечной массы и потере мышечной массы, вызванной возрастным возрастом. -Ассоциированные сопутствующие заболевания. Достижение этой цели, вероятно, будет ускорено за счет использования широкого спектра новых и существующих методологий pari passu.
body_by_mma

Регулирование массы скелетных мышц с помощью упражнений и питания
Прочёл все 25 000 букв, коллега. Вполне читабельный гугл-перевод. Гуглу респект.
Однако, вы всё-таки не забывайте указывать источник. Важно ведь не только оценить исследование, но и прошерстить автора, цепочку цитирования - чтобы комплексно взглянуть на проблему.
По теме вопроса: оно понятно. Проблема всех этих исследований в том, что ОНО (механизм, сценарий) ПОЧЕМУ-ТО на определённом этапе перестаёт работать. НАЧИНАЕТСЯ же у всех всегда всё прекрасно.
Поэтому и НЕ интересны механизмы и сценарии новичков. А интересны как раз механизмы обхода тупика, возникающего в моменте, когда новичковость перетекает в опытность.
----------
Так шта нужно искать ДЛИТЕЛЬНЫЕ исследования об отзывчивости рецепторов на стимуляцию тренингом - на иммунизацию этих рецепторов (снижение чувствительности) и ре-иммунизацию длительными периодами отдыха.
IgG к SARS-CoV-2
IgG к SARS-CoV-2
ВОт этот момент понравился
Хотя мы многое узнали о регуляции обмена мышечного белка из исследований, в которых вводили изолированный белок или аминокислоты, пища - это то, что люди едят, и стало очевидно, что, хотя белок / аминокислоты являются основным анаболическим стимулом для мышц, источником диетический белок и взаимодействие с пищевым матриксом могут иметь важное влияние на биологическую активность белка / аминокислот. В то время как потребление богатого белком постного мяса, как было обнаружено, стимулирует MPS зависимым от дозы криволинейным образом как у молодых, так и у пожилых людей (Symons et al. 2009; Robinson et al. 2013), а реакция MPS на употребление нежирной говядины была максимальной. с размером порции ∼113–170 г (4–6 унций), что обеспечивает ∼30–35 г белка.
Тут мы получили циферку оптимального размера порции мяса
body_by_mma
Поэтому и НЕ интересны механизмы и сценарии новичков. А интересны как раз механизмы обхода тупика, возникающего в моменте, когда новичковость перетекает в опытность.
Ну, собственно: ищем антоним к термину "новичок". "Опытный"? А что если - "старичок"? ("В бой идут одни старики").
А потом смотрим сюда:
Старение приводит к характерным изменениям в обмене мышечного белка, называемым возрастной «анаболической резистентностью» (Moore et al. 2015), которая характеризуется притупленной реакцией как MPS, так и MPB на анаболические эффекты аминокислот и упражнений и антипротеолитический эффект инсулина, приводящий к постепенной потере мышечной массы. Первое свидетельство этого феномена было предоставлено профессором Ренни и его коллегами (Катбертсон и др., 2005; Уилкс и др., 2009; Кумар и др., 2009a), которые продемонстрировали, что, хотя базальные скорости МПС и МПБ не различаются у молодых и у пожилых людей увеличение MPS в результате приема аминокислот уменьшается у пожилых людей. Позже они продемонстрировали, что опосредованное инсулином подавление MPB (Wilkes et al. 2009) и вызванное физическими упражнениями увеличение MPS (Kumar et al. 2009b) также снижаются у пожилых людей по сравнению с молодыми людьми. Наличие анаболической резистентности у пожилых людей с тех пор подтверждено несколькими другими исследователями (Moore et al. 2015). Кроме того, эта анаболическая резистентность больше поражает пожилых женщин, чем мужчин (Smith et al. 2012). Поэтому стратегии профилактики и лечения возрастной саркопении сосредоточены на преодолении этой анаболической резистентности (Bauer et al. 2013).
Вот, собственно, и ответ. На старте спектра мы имеем молодого генетика мужчину, на финише – пожилого нонреспондера женщину.
И с каждым наращенным килограммом мышечной массы человек перемещается всё правее и правее по спектру - пока не замёрзнет в определённой точке. Смайл мышцы молодого, пересаженной старику и восстанавливающейся столь же медленно, как мышца старика - и наоборот.
Анаболическая резистентность возрастает вместе с ростом общего количества мышечной массы, удерживаемой на теле - попросту из-за того, что гормонов на всё НЕ ХВАТАЕТ.
Сообщение изменено: Михалы4 (16 августа 2020 - 01:35)
IgG к SARS-CoV-2
Ну, собственно: ищем антоним к термину "новичок". "Опытный"? А что если - "старичок"? ("В бой идут одни старики").
А потом смотрим сюда:
Вот, собственно, и ответ. На старте спектра мы имеем молодого генетика мужчину, на финише – пожилого нонреспондера женщину.
И с каждым наращенным килограммом мышечной массы человек перемещается всё правее и правее по спектру - пока не замёрзнет в определённой точке. Смайл мышцы молодого, пересаженной старику и восстанавливающейся столь же медленно, как мышца старика - и наоборот.
Анаболическая резистентность возрастает вместе с ростом общего количества мышечной массы, удерживаемой на теле - попросту из-за того, что гормонов на всё НЕ ХВАТАЕТ.
Кстати не факт что гормонов вообще и тестостерона в частности. Т.к. у женщин его в 10 раз меньше.
IgG к SARS-CoV-2
Чтобы максимизировать ресинтез гликогена после тренировки, следует употреблять углеводную добавку, превышающую 1,0 г на кг (-1) массы тела, сразу после соревнований или тренировочного боя. Продолжение приема добавок каждые два часа будет поддерживать высокую скорость накопления до шести часов после тренировки. Добавки, состоящие из глюкозы или полимеров глюкозы, являются наиболее эффективными для восполнения гликогена в мышцах, тогда как фруктоза наиболее полезна для восполнения гликогена в печени. Добавление белка к углеводной добавке может также увеличить скорость хранения гликогена из-за способности белка и углеводов синергетически влиять на секрецию инсулина.
https://pubmed.ncbi....ih.gov/9694422/
ПериодизаторЪ

Добавление белка к углеводной добавке может также увеличить скорость хранения гликогена из-за способности белка и углеводов синергетически влиять на секрецию инсулина.
Так в прошлом веке появились "Гейнеры" ... белково-углеводные смеси в спортивном питании.
Аксакал

Чтобы максимизировать ресинтез гликогена после тренировки
Аксакал

Чтобы максимизировать ресинтез гликогена после тренировки, следует употреблять углеводную добавку, превышающую 1,0 г на кг (-1) массы тела, сразу после соревнований или тренировочного боя. Продолжение приема добавок каждые два часа будет поддерживать высокую скорость накопления до шести часов после тренировки. Добавки, состоящие из глюкозы или полимеров глюкозы, являются наиболее эффективными для восполнения гликогена в мышцах, тогда как фруктоза наиболее полезна для восполнения гликогена в печени. Добавление белка к углеводной добавке может также увеличить скорость хранения гликогена из-за способности белка и углеводов синергетически влиять на секрецию инсулина.
На кой хрен вот эта инфа качку? Актуальна может быть для спортсменов (в большей степени цикликов) в соревновательный сезон между стартами (допустим, как в лыжах, биатлоне). А в планомерном тренировочном процессе в межсезонье эта инфа и им ограниченно важна или вообще побоку до определённого периода.
ОколоКроссфит

Сырые чуть ли не вдвое хуже усваиваются. Высрешь половину ))
 К еде я не привередлив и могу жрать многое, в том числе и в сыром виде
К еде я не привередлив и могу жрать многое, в том числе и в сыром виде
0 пользователей, 17 гостей, 0 скрытых
 Наверх
Наверх