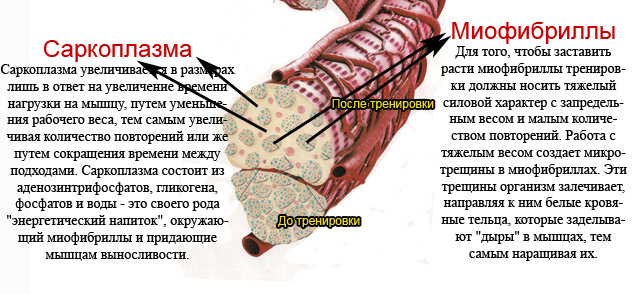
Михалыч, в видео для борцов про метод тренировки МХ то, что ты говоришь относится только к циклическим видам спорта.
Да неужели? Причём тут вообще "вид спорта"??
То есть, если нога давит на педаль велосипеда – метод работает, а если на платформу жима ногами – то уже нет?
Для НЕцикличесих видов спорта им предлагается следующий метод тренировки МХ в глиполиптических МВ - 3-5 сек высокоинтенсивной более 80% нагрузки (бег например, если для ног), а затем отдых 15-25 с (правило для определения отдыха у него - время нагрузки умножается на 5, смотрим с 7мин 30с), количество сессий за тренировку 10-40, частота тренировок в неделю 2-7. Такие трени одновременно ведут и к росту МФ в БМВ.
Ну и какой же из всего этого следует вывод?
Ты сам-то хоть разобрался с механизмом развития митохондрий?
Сложные схемы, как разновидность простых, действенность последних отнюдь не отменяют.
А если разобраться – то лишь подтверждают.
Ты сам-то хоть разобрался с механизмом развития митохондрий?
Ответь на несколько вопросов:
1. Что является стрессом для митохондрий (МХ)?
2. Найди этот стресс-фактор в специфическом протоколе нагрузки (СПН), предлагаемом Селуяновым для «МХ + МФ БМВ».
3. Чем обоснован период отдыха между сетами в предлагаемом СПН?
А тренировка на МХ и МФ в ММВ у Селуянова совпадают.
Что там и с чем "совпадает"??
Точно так же, как и в вышеприведённом варианте для БМВ, предложен комплексный метод тренировки, развивающий и миофибриллы (МФ) и митохондрии (МХ) в ММВ. Но это вовсе не означает, что стимул для МХ – такой же, как и стимул для МФ.
Можно, при желании, тренировать МХ ММВ отдельно, и МФ ММВ – отдельно, и ничего там совпадать не будет.
Ответь на те же вопросы, что и по СПН «МХ + МФ БМВ» (см. выше), и всё встанет на свои места.
ММВ растут максимум процентов на 10 в поперечнике. Забейте...
Не согласен.
ММВ также как и БМВ гипертрофируют примерно вдвое.
Другое дело, что они изначально тоньше чем БМВ примерно в те же два раза, то есть гипертрофированное ММВ по толщине равно нетренированному БМВ.
Однако, если учесть, что их пул весьма велик, то...
И, опять же, повторю то, что высказывал уже не раз – ЛЮБАЯ тренировка, направленная на гипертофию БМВ развивает в равной мере и ММВ. Все попытки развивать ММВ отдельно считаю не более чем спекуляцией околоспортивных шарлатанов навроде Селуянова.
очевидно в разных волокнах с разной скоростью
Максим, то что ты пишешь про какие-то там "разные" волокна не имеет НИКАКОГО отношения к обсуждаемому вопросу. Я уже
писал тебе об этом, не знаю, прочитал ты уже это сообщение, или ещё нет...
Сообщение изменено: Михалы4 (11 апреля 2012 - 01:06)
 Вход
Вход Регистрация
Регистрация












 Наверх
Наверх


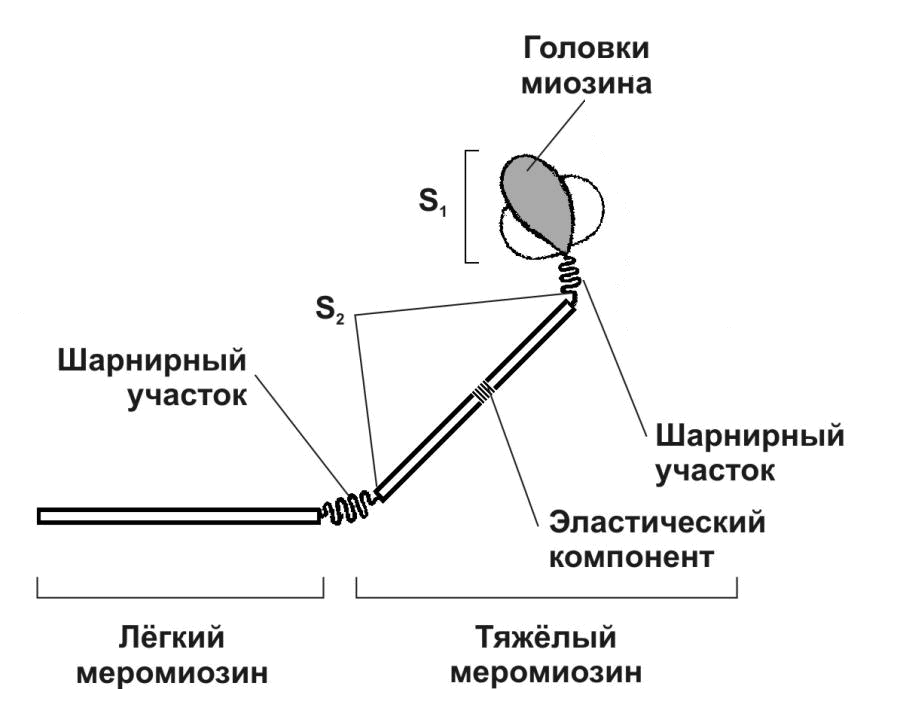



 Это "вода в трубе" - в один конец трубы втекает, из другого вытекает - но в этой трубе постоянно находится одинаковый объём воды. Растет давление, увеличивается скорость - пох, объем один и тот же, пока трубу не передавишь со стороны подачи. Если не будет запасов "воды" - "труба" высохнет, поэтому организм в первую очередь восполняет наиболее быстро мобилизуемые источники энергии - КрФ. Всего 1 молекула креатинфосфата, всего одна химическая реакция = 1 АТФ. Огромное преимущество по сравнению с анаэробным и аэробным гликолизом в скорости подачи энергии. Поэтому первые же свободные АТФ идут на синтез КрФ.
Это "вода в трубе" - в один конец трубы втекает, из другого вытекает - но в этой трубе постоянно находится одинаковый объём воды. Растет давление, увеличивается скорость - пох, объем один и тот же, пока трубу не передавишь со стороны подачи. Если не будет запасов "воды" - "труба" высохнет, поэтому организм в первую очередь восполняет наиболее быстро мобилизуемые источники энергии - КрФ. Всего 1 молекула креатинфосфата, всего одна химическая реакция = 1 АТФ. Огромное преимущество по сравнению с анаэробным и аэробным гликолизом в скорости подачи энергии. Поэтому первые же свободные АТФ идут на синтез КрФ.






 ) в организме.
) в организме.